发生遗传学(发生遗传学)

正文
发生遗传学
从遗传学观点看来发育是从基因型转化为表型的过程。从受精开始,以至胚层、器官原基的形成,组织、细胞的决定和分化,每一步都要受特定基因的控制。这些基因发生突变就会相应地造成发育的异常、停顿、甚至胚胎的死亡。影响因素
发生遗传学在方法学上主要是利用这些影响发育的突变型,并结合实验胚胎学、细胞生物学和分子生物学的方法,从不同水平来分析基因和性状发育之间的关系,以阐明基因控制发育的机理。因此它同胚胎学、畸胎学、细胞生物学和分子生物学,尤其是与细胞分化过程中基因表达及其调控方面的研究有着密切的关系。
简史
遗传是发育的基础遗传是发育的基础,而发育是遗传的实现。两者之间的关系历来是遗传学家和胚胎学家共同关心的问题。早在19世纪末期,德国生物学家A.魏斯曼就曾经试图建立发育和遗传的统一理论。他曾经假定全部发育过程是受细胞核控制的,卵裂过程中核内遗传物质的不等分配是胚胎分化的主要原因。这一理论的原有形式虽然被早期实验胚胎学的实验事实所否定,但他提出的问题却吸引了几代生物学家的继续探讨。美国细胞学家E.B.威尔逊1928年在《发育和遗传中的细胞》一书中提出基因在细胞水平的活动是发育的根本原因这一论点,认为发育是“遗传特性按一定时、空秩序的表现”。基因论的创建人美国遗传学家兼实验胚胎学家T.H.摩尔根在他晚年(1934)的著作《胚胎学和遗传学》一书中也强调遗传学和胚胎学统一的重要性,并提出在发育的“不同时期有不同的一组基因起作用”的论点。
最早用实验方法最早用实验方法确定了染色体在发育中的重要性的是德国实验胚胎学家T.H.博韦里。他根据对海胆受精卵分裂球发育的分析结果,认为正常发育依赖全套染色体的正常组合,每一个染色体对发育都有特殊的影响。美国遗传学家R.B.戈德施米特对基因在发育中的作用给以很大的重视。他在1935年发现拟表型,认为基因突变和环境因子的作用一样,都可能干扰相同的发育过程。致死基因和畸型发育的研究对发生遗传学的形成也起过重要的推动作用。
1955年瑞士实验胚胎学家E.哈多恩在《发生遗传学和致死因子》一书中系统地总结了致死突变型引起异常发育的大量材料,并用实验胚胎学方法对这些病理发育过程进行分析,促进了对正常发育中基因作用机制的了解。1963年H.格吕内贝格在《发育病理学》一书中提出基因作用多效性概念,认为胚胎是一个高度复杂的相互作用系统,任何与发育有关的突变都可能对胚胎发育产生广泛的影响。因此应透过对各种病理发育现象的分析找出基因在细胞水平作用的原初效应。70年代初 C.L.马克特和 H.乌尔施普龙合写了《发生遗传学》,把当时散见于各方面的有关资料汇总起来,初创了这门学科的体系。
生理原因长期以来胚胎学家致力于用实验方法分析发育的生理原因,尤其着重组织者的研究,因而忽视了发育的遗传基础。对异属诱导系统的实验结果说明诱导者只起激发作用,被诱导出来的器官的种属特性则取决于起反应的细胞本身的遗传特性。例如把蛙的外胚层移植到蝾螈的头部,被诱导形成的口器是蛙特有的角质腭和吸盘。这些事实促使胚胎学家注意发育和遗传关系问题。1940年英国实验胚胎学家兼遗传学家C.H.沃丁顿在《组织者和基因》一书中首先提出从基因和细胞质环境的相互作用来理解反应能力、诱导和决定等胚胎学基本概念的主张。但限于当时生物学发展水平,还不能对基因控制发育的分子机制作深入的研究。
50年代中期以来的分子生物学的重大进展使解决遗传和发育关系问题的条件逐渐成熟起来。遗传信息传递的中心法则揭示了生物的遗传和发育的内在联系。从分子水平看来,细胞分化和性状发育都是表型专一的大分子合成的结果,因而归根结蒂依赖基因在发育过程中按一定的时空秩序的表达。基因的表达又可以用相应的信使核糖核酸(mRNA)的转录和专一的蛋白质(如结构蛋白、酶等)的合成来追踪。1976年美国分子生物学家E.戴维森所写的《早期发育中的基因活动》一书代表从分子水平探讨发育和遗传关系问题的发展趋向。目前研究核酸分子结构和功能的方法日臻完善,尤其是遗传工程技术提供了前所未有的研究基因结构、表达及其调控的有力的手段。这就为从分子水平探讨发生遗传学问题,特别是研究影响特定发育过程的单个基因(或基因群)的作用开辟了新的前景。
对象和方法
发生遗传学开始于对黑腹果蝇发育的遗传学分析。从摩尔根时代以来已积累了大量有关果蝇遗传和发育的实验资料和许多影响发育的突变型。果蝇的唾腺巨大染色体上疏松区的消长可以反映不同发育时期的基因活动情况。此外,对体表形态构造的变异也便于作精细的描述。这些都是果蝇这一实验材料的优点,缺点是较难得到足够量的可供生化分析的样品。另一个常用的材料是小鼠,也有许多遗传背景清楚的纯系和致死突变型(如T基因座位)可供发生遗传学研究,不过同样存在取材上的困难。用海胆作为研究对象则便于取得较大量的材料,特别是同一发育阶段的材料,可是可供研究的突变型较少。英国分子生物学家 S.布伦纳在 60年代倡导用一种营自由生活而繁殖迅速的秀丽隐杆线虫(Caenorabditis elegans) 作材料。这种动物身体和各器官的细胞数少而恒定,胚胎发育属于镶嵌型,每一个器官的来源有确定的细胞谱系可循,而且便于用实验方法得到许多突变型。低等真核生物如粘菌(Dictyosteliumdiscoideum)由于生活史和形态发生的特点,也被一些人用来作为发生遗传学的研究对象。此外,甚至有人采用更为简单的对象如细菌形成芽孢的过程、噬菌体的自动装配过程等作为一个发育模型来进行发生遗传学研究。
应用突变型进行遗传学分析是发生遗传学的基本研究方法。此外,实验胚胎学方法(如移植、离体培养等)、细胞生物学方法(如核移植、细胞化学技术等)、生化方法(如同功酶测定等)和分子生物学方法(如重组DNA技术、分子杂交等)都是常用的实验手段。70年代以来重组DNA技术的应用使有可能直接研究没有发生突变的野生型基因的作用。
研究主题
动物的发育是从受精开始,通过受精卵的核质之间、分裂球之间、以及胚胎不同部位之间的相互作用,使基因按一定时空秩序表达,从而控制细胞和器官原基的逐步决定和分化的过程。动物的发育类型(镶嵌卵或调整卵)不同,基因控制发育的具体方式也可能不同。果蝇(镶嵌卵)的发育中细胞的发育命运决定得早,胚胎发育主要表现为一连串由大到小的发育区逐步划分的过程。两栖类(调整卵)的发育中细胞的命运决定得晚,细胞迁移和相互作用在发育中起重要的作用。截至20世纪70年代发生遗传学的知识还主要是从果蝇上得到的。
核质关系和母体效应
决定因素细胞核移植实验证明果蝇前囊胚层细胞核具有全部发育潜能,而囊胚层细胞核的发育潜能已开始受到限制,这可能是受到卵表层细胞质的影响的结果。紫外线损伤和卵质移植实验证明位于卵后端的极质能决定迁移到这区域中的细胞核的发育命运,使它们形成原始生殖细胞。无尾两栖类(如蛙和瓜蟾)中也存在类似的现象,它们的原始生殖细胞是由位于卵的植物极的生殖质决定的。
极质区域绝孙突变型雌性果蝇的囊胚细胞核不能及时迁入极质区域,生殖细胞因而不能正常发生。这表明卵质特性是在卵子发生过程中受母体基因决定的,这一现象称为母体效应。某些母体效应突变型如双腹端(bicaudal,bic)和背方 (dorsal,dl)能广泛影响胚胎发育的格局。纯合双腹端 (bic/bic)雌蝇所产的一部分卵只发生成为两个腹端,彼此沿前后轴对称排列,而缺少头、胸和其他腹节。纯合背方 (dl/dl)雌蝇的胚胎构造全部背方化,而前后极性仍然保持。这两个母体效应基因各自定位在染色体的一个位置上。有人假定它们的野生型基因产物的浓度在卵内各自沿前后轴和背腹轴呈梯度分布,从而决定了果蝇胚胎的正常发育格局。
细胞的决定和发育区划分
定义利用带有不同遗传标记的胚胎细胞的嵌合体可以追溯细胞决定的时期、发育命运以及器官原基奠基细胞的数目。得到嵌合体的常用方法有3种:
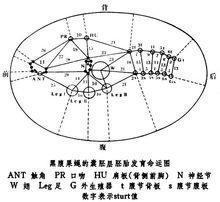
雌雄嵌合体在果蝇中可以利用环状X染色体常在受精卵的第一次有丝分裂过程中丢失的特性取得雌雄嵌合体。例如雌性亲本的两个 X染色体上全部都是野生型基因,不过其中一个是环状染色体;雄性亲本的一个X染色体上带有3个隐性突变基因(白眼w,黄体y和分叉刚毛bi)。这两个亲本交配后可以产生一种带有一个环状X染色体和一个有3个隐性基因的X染色体的受精卵。这种受精卵第一次卵裂核分裂后,一个子细胞带有这两个X染色体,因而发育成嵌合体上具有野生型表型的雌性部分(XX);另一个子细胞由于丢失了环状 X染色体,因而发育成为具有突变型表型的雄性部分(X0)。两部分之间有清楚的分界面。由于第一次核分裂的纺锤体的取向不同,含XX和X0细胞群的分界面也有不同。因此所得到雌雄嵌合体可以是左右各半、前后各半或者是其他种种形式。利用雌雄嵌合体的这些特点,便可以制作各器官原基在囊胚层上的预定命运图。其原理是两个成体器官(如翅和头)的原基细胞在囊胚层上相距愈近,则嵌合体分界线通过两原基之间的机率就愈小。因此带有不同遗传标记的两个构造在嵌合体上出现的百分比可作为这两个原基之间相对距离的量度(为了纪念首先提出这种设想的A.H.斯特蒂文特,用sturt作为单位,一个sturt代表 1%个体中嵌合体分界线通过所研究的两个原基)。如果再测出这两个构造中每一个对第三个构造(如吻)的相对距离就可以确定某一器官的原基在囊胚层上的相对位置。推而广之,便可以做出各器官在囊胚层上的预定命运图(见图)。
重组嵌合体在一定发育时期用X射线诱发胚胎细胞中的有丝分裂交换(见连锁和交换),使单个胚胎细胞及其子细胞群带上遗传标记,便可鉴定发育时期细胞的决定状态。例如用X射线照射小体(M)杂合体(M/M+)果蝇的囊胚层期胚胎,可以诱发体细胞染色体交换而在生长缓慢的杂合体细胞背景上出现一片生长迅速的野生型纯合M+/M+细胞。这种细胞群决不跨越两个体节,说明体节的决定可能发生在囊胚层期或稍后的一次分裂期。晚一些时候照射所得到的一个标记细胞群决不越过同一体节的前部和后部的分界线,说明体节的前部和后部这时已经决定。再晚些时候照射,得到的一个标记细胞群就不再同时包含背方和腹方的构造,也就是说发生了背腹的分区。器官发生过程中还会逐渐发生更进一步的分离。象这样每一来源于少数几个奠基细胞的细胞群所占据身体或器官上的一定区域称为发育区。属于一个发育区的细胞决不会越界同其他发育区的细胞混合。发育区可看作是发育的基本单位,它在许多方面具有“场区”的性质。同一发育区内发育格局可以调整,而不同发育区之间则是镶嵌的、不可调整的。果蝇的发育过程是一系列由大到小发育区愈分愈细的过程。这一现象称为发育区划分。
异表型嵌合体把基因型不同的两种小鼠的早期胚胎在体外人工地并合在一起,然后移回到母鼠子宫内,便能发育成嵌合体,称为异表型嵌合体。通过异表型嵌合体研究,在小鼠中测定了色素细胞、毛囊、生殖腺以及其他内脏的奠基细胞的数目。例如生殖腺的奠基细胞数是2~9。
发育途径的决定和转变
果蝇成虫的器官原基果蝇成虫的器官原基──成虫盘在胚胎发育的早期已经决定了,但要等到幼虫变态时受到激素的影响才开始分化。E.哈多恩曾把成虫盘(如生殖板原基)移植到成虫腹腔内,经过连续70次以上的传代(超过1000次细胞分裂)后再移植到将要变态的幼虫体内仍然可以按照预定的命运分化为生殖板。这说明细胞的决定和分化在时间上是可以分隔的。细胞虽然经过了上千次的分裂仍然可以保持原来的决定状态。然而成虫盘经过若干次移植传代后偶尔可能改变其决定状态,分化成和它原来的预定命运不同的器官(如触角)。这一现象称为转决,它说明细胞的发育命运可以改变。
同源异形突变型影响果蝇发育途径的还有一种突变型称为同源异形突变型,它能使一种发育途径转变到另一种发育途径,例如使平衡棒原基发育成为翅,触角原基发育成为足等。同源转化的影响限于一个发育区内,如反双胸突变(cbx)能将中胸后部(翅的后部)转化为后胸后部(平衡棒的后部)。根据同源异形突变型的作用方式,有人设想它的野生型基因在正常发育过程中可能起选择和维持特定发育途径的作用,因此称之为选择基因。果蝇胚胎发育中发育区的划分是由大到小逐步进行的,其中每一步都可能受一个(或一组)选择基因的控制。发育程序的遗传控制
时间和地位的基因在果蝇和小鼠等动物中都发现有一类控制某些酶活性在胚胎发育中出现的时间和地位的基因,称为时序基因。例如黑腹果蝇的淀粉酶为2号染色体上两个头尾相连的结构基因所编码。在它们的附近有一个基因map,它的3个等位基因mapa、mapb、mapc控制着淀粉酶在中肠出现的时间和地位。小鼠的 5号染色体上有一个编码β-葡萄糖苷酸酶的结构基因Gus,和它紧密连锁的有一个控制酶出现时间的时序基因Gut。Gut基因发生突变后,β-葡萄糖苷酸酶在各种组织中出现的时间便被扰乱。
幼虫唾腺染色体此外,果蝇等双翅目昆虫的幼虫唾腺染色体上的疏松区也是研究基因活动的时空秩序的好材料。疏松区是光学显微镜下可观察到的染色体的局部膨大的部分,累积大量新合成的mRNA,是进行活跃的转录活动的基因所在的地方。果蝇发育过程中染色体不同部位的疏松区按一定的时间顺序出现,说明这些基因在不同的发育时期起作用。蜕皮激素能诱发特定的疏松区出现,说明这一激素是通过调节该基因的活动而参与控制发育的。
致死突变型致死突变型也常用作研究发育程序的遗传控制的材料,因为使胚胎发育在特定阶段停顿的基因中间有许多是控制该特定发育阶段的基因。许多这类突变型是温度敏感的,在果蝇和线虫都获得了一些温度敏感的突变型,而且已发现不同的突变型常使发育停顿在不同的阶段,某些突变型还影响几个发育阶段。
基因影响发育的机制
基因产物影响发育的基因究竟通过什么途径或基因产物起作用,这也是必须回答的问题。这方面研究得较为深入的是小鼠的T复合座位或T基因复合体,它位于17号染色体上,包括6个互补群。除野生型基因和显性的突变型T以外,还包括一系列的隐性致死或半致死突变型t。杂合体(T/t)具有短尾,纯合体(T/T)则造成胚胎早期畸形和夭折。一系列的隐性致死或半致死突变型 (t)各自对早期发育的一定阶段专一地起作用。例如t12影响滋养外胚层和内胚层细胞团的分离而使发育停顿在桑椹期;t0作用在胚外外胚层和外中胚层分离的时期;t9影响原条期而使中胚层细胞向内迁移受阻,因而造成中轴器官发育异常。进一步分析发现基因的原初效应可能影响细胞表面抗原和它的下方的微丝,从而干扰了原肠运动正常进行。
T 基因座位各个野生型基因的功能可能是控制早期发育中细胞表面特定分子按顺序的表达,从而保证形态发生运动和细胞间相互作用按正常的发育程序有条不紊地进行。对于多数控制发育的基因来说,它们究竟编码什么蛋白质还不清楚,那些控制基因活动的时空秩序的基因的作用机制则更不了解了。
控制发育从分子水平研究基因如何控制发育,对于解决发育和遗传关系这一生物学基本问题有重要的理论意义。在实践上,这方面研究的进展又是了解病理发育(畸胎、肿瘤等)的发生机制,以及把遗传工程方法应用到高等动物,治疗遗传病或改变经济动物遗传性的必要的理论前提。


