蛋白质合成(碱基排列顺序变为蛋白质的过程)

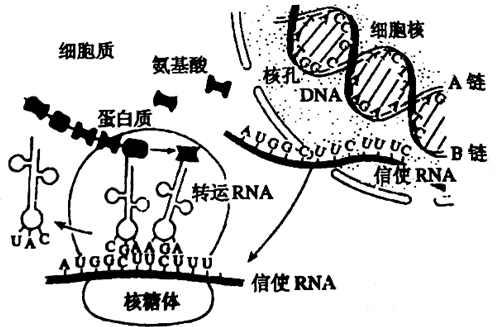
蛋白质合成是指生物按照从脱氧核糖核酸(DNA)转录得到的信使核糖核酸(mRNA)上的遗传信息合成蛋白质的过程。蛋白质生物合成亦称为翻译(Translation),即把mRNA分子中碱基排列顺序转变为蛋白质或多肽链中的氨基酸排列顺序过程。 这是基因表达的第二步,产生基因产物蛋白质的最后阶段。不同的组织细胞具有不同的生理功能,是因为它们表达不同的基因,产生具有特殊功能的蛋白质,参与蛋白质生物合成的成份至少有200种,其主要体是由mRNA、tRNA、核糖核蛋白体以及有关的酶和蛋白质因子共同组成。
蛋白质合成
Protein Synthesis
分子生物学
碱基排列顺序转变为蛋白质
核糖体
mRNA、tRNA
在mRNA的开放式阅读框架区
不同mRNA序列的分子大小
合成过程
蛋白质合成
原核生物与真核生物的蛋白质合成过程中有很多的区别,真核生物此过程更复杂,下面着重介绍原核生物蛋白质合成的过程,并指出真核生物与其不同之处。蛋白质生物合成可分为五个阶段,氨基酸的活化、多肽链合成的起始、肽链的延长、肽链的终止和释放、蛋白质合成后的加工修饰。直接模板
翻译模板protein biosynthesis
不同mRNA序列的分子大小和碱基排列顺序各不相同,但都具有5ˊ-端非翻译区、开放阅读框架区、和3ˊ-端非翻译区;真核生物的mRNA的5ˊ-端还有帽子结构、3ˊ-端有长度不一的多聚腺苷酸(polyA)尾。帽子结构能与帽子结合,在翻译时参与mRNA在核糖体上的定位结合,启动蛋白质生物的合成;帽子结构和ployA尾的作用还有稳定RNA;开放阅读框架区与编码蛋白质的基因序列相对应。
遗传密码表在mRNA的开放式阅读框架区,以每3个相邻的核苷酸为一组,代表一种氨基酸或其他信息,这种三联体形势称为密码子(codon)。如图,通常的开放式阅读框架区包含500个以上的密码子。
遗传密码的特点一方向性:密码子及组成密码子的各碱基在mRNA序列中的排列具有方向性(direction),翻译时的阅读方向只能是5ˊ→3ˊ。
二连续性:mRNA序列上的各个密码子及密码子的各碱基是连续排列的,密码子及密码子的各个碱基之间没有间隔,每个碱基只读一次,不重叠阅读。
三简并性:一种氨基酸可具有两个或两个以上的密码子为其编码。遗传密码表中显示,每个氨基酸都有2,3,4或6个密码子为其编码(除甲硫氨酸只有一个外),但每种密码子只对应一个氨基酸,或对应终止信息。
四通用性:生物界的所有生物,几乎都通用这一套密码子表
五摆动性:tRNA的最后一位,和mRNA的对应不完全,导致了简并性
合成场所
核糖体就像一个小的可移动的工厂,沿着mRNA这一模板,不断向前迅速合成肽链。氨基酰tRNA以一种极大的速率进入核糖体,将氨基酸转到肽链上,又从另外的位置被排出核糖体,延伸因子也不断地和核糖体结合和解离。核糖体和附加因子一道为蛋白质合成的每一步骤提供了活性区域。
有关信息
氨基酸活化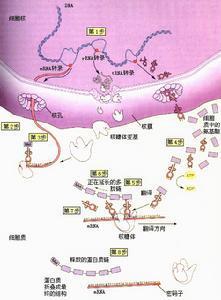
蛋白质合成
在进行合成多肽链之前,必须先经过活化,然后再与其特异的tRNA结合,带到mRNA相应的位置上,这个过程靠tRNA合成酶催化,此酶催化特定的氨基酸与特异的tRNA相结合,生成各种氨基酰tRNA.每种氨基酸都靠其特有合成酶催化,使之和相对应的tRNA结合,在氨基酰tRNA合成酶催化下,利用ATP供能,在氨基酸羧基上进行活化,形成氨基酰-AMP,再与氨基酰tRNA合成酶结合形成三联复合物,此复合物再与特异的tRNA作用,将氨基酰转移到tRNA的氨基酸臂(即3'-末端CCA-OH)上(图1)。简介内容原核细胞中起始氨基酸活化后,还要甲酰化,形成甲酰蛋氨酸tRNA,由N10甲酰四氢叶酸提供甲酰基。而真核细胞没有此过程。前面讲过运载同一种氨基酸的一组不同tRNA称为同功tRNA。一组同功tRNA由同一种氨酰基tRNA合成酶催化。氨基酰tRNA合成酶对tRNA和氨基酸两者具有专一性,它对氨基酸的识别特异性很高,而对tRNA识别的特异性较低。氨基酰tRNA合成酶是如何选择正确的氨基酸和tRNA呢?按照一般原理,酶和底物的正确结合是由二者相嵌的几何形状所决定的,只有适合的氨基酸和适合的tRNA进入合成酶的相应位点,才能合成正确的氨酰基tRNA。现在已经知道合成酶与L形tRNA的内侧面结合,结合点包括接近臂,DHU臂和反密码子臂(图2)。氨基酰-tRNA合成酶与tRNA的相互作用,可见氨酸接受柄、乍看起来,反密码子似乎应该与氨基酸的正确负载有关,对于某些tRNA也确实如此,然而对于大多数tRNA来说,情况并非如此,人们早就知道,当某些tRNA上的反密码子突变后,但它们所携带的氨工酸却没有改变。1988年,候稚明和Schimmel的实验证明丙氨酸tRNA酸分子的氨基酸臂上G3:U70这两个碱基发生突变时则影响到丙氨酰tRNA合成酶的正确识别,说明G3:U70是丙氨酸tRNA分子决定其本质的主要因素。tRNA分子上决定其携带氨基酸的区域叫做副密码子。一种氨基酰tRNA合成酶可以识别以一组同功tRNA,这说明它们具有共同特征。例如三种丙氨酸tRNA(tRNAAlm/CUA,tRNAAim/GGC,tRNAAin/UGC都具有G3:U70副密码子。)但没有充分的证据说明其它氨基酰tRNA合成酶也识别同功tRNA组中相同的副密码子。另外副密码子也没有固定的位置,也可能并不止一个碱基对。
多肽链解析
核蛋白体大小亚基,mRNA起始tRNA和起始因子共同参与肽链合成的起始。
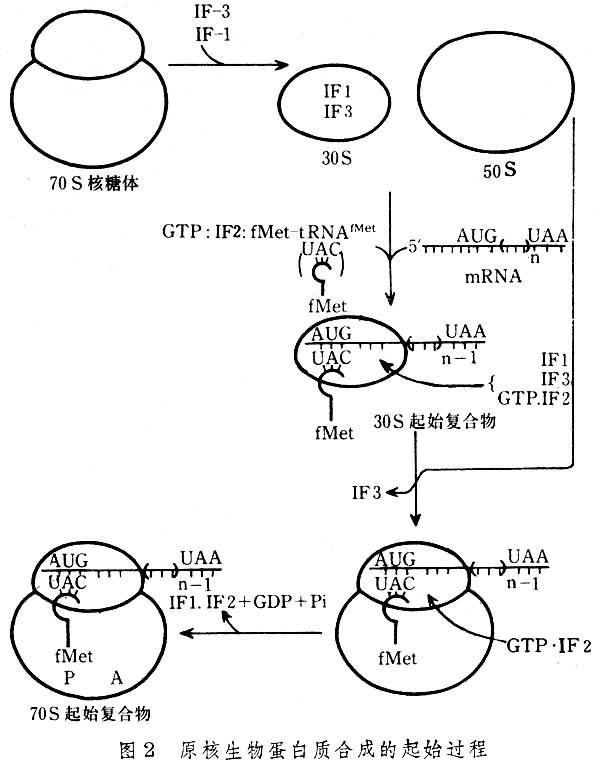
1、大肠杆菌细胞翻译起始复合物形成的过程:
⑴核糖体30S小亚基附着于mRNA起始信号部位:原核生物中每一个mRNA都具有其核糖体结合位点,它是位于AUG上游8-13个核苷酸处的一个短片段叫做SD序列。这段序列正好与30S小亚基中的16S rRNA3’端一部分序列互补,因此SD序列也叫做核糖体结合序列,这种互补就意味着核糖体能选择mRNA上AUG的正确位置来起始肽链的合成,该结合反应由起始因子3(IF-3)介导,另外IF-1促进IF-3与小亚基的结合,故先形成IF3-30S亚基-mRNA三元复合物。
⑵30S前起始复合物的形成:在起始因子2作用下,甲酰蛋氨酰起 始tRNA与mRNA分子中的AUG相结合,即密码子与反密码子配对,同时IF3从三元复合物中脱落,形成30S前起始复合物,即IF2-3S亚基-mRNA-fMet-tRNAfmet复合物,此步需要GTP和Mg2+参与。
蛋白质合成
⑶70S起始复合物的形成:50S亚基上述的30S前起始复合物结合,同时IF2脱落,形成70S起始复合物,即30S亚基-mRNA-50S亚基-mRNA-fMet-tRNAfmet复合物。此时fMet-tRNAfmet占据着50S亚基的肽酰位。而A位则空着有待于对应mRNA中第二个密码的相应氨基酰tRNA进入,从而进入延长阶段,以上过程见图3和图4。2、真核细胞蛋白质合成的起始真核细胞蛋白质合成起始复合物的形成中需要更多的起始因子参与,因此起始过程也更复杂。
⑴需要特异的起始tRNA即,-tRNAfmet,并且不需要N端甲酰化。已发现的真核起始因子有近10种(eukaryote Initiation factor,eIF)
⑵起始复合物形成在mRNA5’端AUG上游的帽子结构,(除某些病毒mRNA外)
⑶ATP水解为ADP供给mRNA结合所需要的能量。
真核细胞起始复合物的形成过程是:
翻译起始也是由eIF-3结合在40S小亚基上而促进80S核糖体解离出60S大亚基开始,同时eIF-2在辅eIF-2作用下,与Met-tRNAfmet及GTP结合,再通过eIF-3及eIF-4C的作用,先结合到40S小亚基,然后再与mRNA结合。mRNA结合到40S小亚基时,除了eIF-3参加外,还需要eIF-1、eIF-4A及eIF-4B并由ATP水解为ADP及Pi来供能,通过帽结合因子与mRNA的帽结合而转移到小亚基上。但是在mRNA5’端并未发现能与小亚基18SRNA配对的S-D序列。目前认为通过帽结合后,mRNA在小亚基上向下游移动而进行扫描,可使mRNA上的起始密码AUG在Met-tRNAfmet的反密码位置固定下来,进行翻译起始。
肽链步骤
多肽链的延长在多肽链上每增加一个氨基酸都需要经过进位,转肽和移位三个步骤。⑴为密码子所特定的氨基酸tRNA结合到核蛋白体的A位,称为进位。氨基酰tRNA在进位前需要有三种延长因子的作用,即,热不稳定的E(Unstable temperature,EF)EF-Tu,热稳定的EF(stable temperature EF,EF-Ts)以及依赖GTP的转位因子。EF-Tu首先与GTP结合,然后再与氨基酰tRNA结合成三元复合物,这样的三元复合物才能进入A位。此时GTP水解成GDP,EF-Tu和GDP与结合在A位上的氨基酰tRNA分离。
多肽链详情
一级结构加工修饰⑴N端甲酰蛋氨酸或蛋氨酸的切除:N端甲酰蛋氨酸是多肽链合成的起始氨基酸,必须在多肽链折迭成一定的空间结构之前被切除。其过程是:① 去甲酰化;② 去蛋氨酰基。
⑵氨基酸的修饰:由专一性的酶催化进行修饰,包括糖基化、羟基化、磷酸化、甲酰化等。
⑶二硫键的形成:由专一性的氧化酶催化,将-SH氧化为-S-S-。
⑷肽段的切除:由专一性的蛋白酶催化,将部分肽段切除。
高级结构的形成⑴构象的形成:在分子内伴侣、辅助酶及分子伴侣的协助下,形成特定的空间构象。
⑵亚基的聚合。⑶辅基的连接。
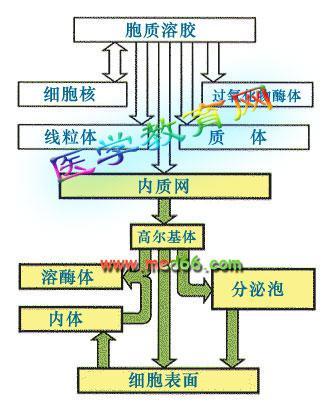
靶向输送蛋白质合成后,定向地被输送到其执行功能的场所称为靶向输送。大多数情况下,被输送的蛋白质分子需穿过膜性结构,才能到达特定的地点。因此,在这些蛋白质分子的氨基端,一般都带有一段疏水的肽段,称为信号肽。分泌型蛋白质的定向输送,就是靠信号肽与胞浆中的信号肽识别粒子(SRP)识别并特异结合,然后再通过SRP与膜上的对接蛋白(DP)识别并结合后,将所携带的蛋白质送出细胞。
信号肽假说:信号肽位于新合成的分泌蛋白N端。对分泌蛋白的靶向运输起决定作用。①细胞内的信号肽识别颗粒(SRP)识别信号肽,使肽链合成暂时停止,SRP引导核蛋白体结合粗面内质网膜;②SRP识别、结合内质网膜上的对接蛋白,水解GTP使SRP分离,多肽链继续延长;③信号肽引导延长多肽进入内质网腔后,经信号肽酶切除。分泌蛋白在高尔基体包装成分泌颗粒出胞。
生物调控
蛋白质合成的调控生物体内蛋白质合成的速度,主要在转录水平上,其次在翻译过程中进行调节控制。它受性别、激素、细胞周期、生长发育、健康状况和生存环境等多种因素及参与蛋白质合成的众多的生化物质变化的影响。由于原核生物的翻译与转录通常是偶联在一起的,且其mRNA的寿命短,因而蛋白质合成的速度主要由转录的速度决定。弱化作用是一个通过翻译产物的过量与不足首先影响转录,从而调节翻译速度的一种方式。mRNA的结构和性质也能调节蛋白质合成的速度。
HCR两种状态真核生物转录与翻译不是偶联的,通常蛋白质合成的速度比原核生物慢。真核生物除了主要通过转录和转录后加工及mRNA的结构和性质(如帽子结构和多聚A尾巴等)(见信使核糖核酸)进行调控外,通过对珠蛋白生物合成研究表明,真核起始因子eIF-2是翻译速度的限制因子,因此影响eIF-2的因素能调节翻译的速度。用哺乳动物网织红细胞的无细胞制剂进行离体研究指出,当缺乏血红素时,因为无法形成血红蛋白,没有必要合成蛋白质。实验证明血红素的调控是通过一种称为血红素调控阻遏物(HCR)实现的。HCR有活泼和不活泼的两种状
血红素的影响血红素通过影响eIF-2对蛋白质进行调控。当血红素存在时,抑制了胞蛋白质合成,而且还能促进通常不合成血红蛋白的细胞合成蛋白质,如促进肝癌细胞、海拉细胞和腹水瘤细胞无细胞制剂的蛋白质合成。
蛋白质抑制剂蛋白质生物合成的抑制剂 许多蛋白质生物合成抑制剂具有高度专一性,这对于研究合成机制很重要。许多临床有效的抗生素是通过特异抑制原核生物的蛋白质合成而发挥作用的,它们抑制细菌生长而不损害人体细胞。利用两类生物蛋白质合成的差异,可以找出治疗细菌感染引起的疾病的药物。表中列出一些较为重要的蛋白质生物合成抑制剂及其作用部位和专一性。


