木霉属(木霉属)

VLoG
次浏览
更新时间:2023-05-19
木霉属
基本信息
| 拉丁学名 | Trichoderma |
| 门 | 真菌门 |
| 亚门 | 半知菌亚门 |
| 纲 | 丝孢纲 |
形态特征
1. 木霉属的描述
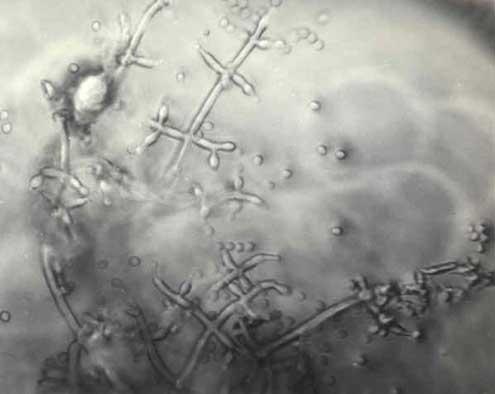
木霉属
分生孢子在平展产孢区或者在平展的产孢簇中产生,产孢簇也可以是紧密结构,开始为白色,后来也可以是白色,但较多的情况是转变为绿色、灰色或者棕色;常常形成厚壁孢子,在基内菌丝上产量大,间生,或者在营养菌丝侧枝的尖端端生,圆形或者椭球形,无色至浅黄色或者绿色,表面光滑或有加厚现象。
分生孢子梗在多数种类中有粗直或者为锯齿状的主轴,初级分枝间距规则,有二级分枝,二级分枝也可以再次分枝,并且有连续的再次分枝,随着分枝次数的增加,在顶端或者远端变短并变细,分枝近似二歧式,或者单生、对生或者轮生。某些种通过重复的轮状分枝形成高度分枝的金字塔形结构。在其他一些种类中,分枝不规则,单生或者对生,而且不再重复分枝。另外一些种的分生孢子梗主轴和初级分枝常常以不育的分生孢子梗结束,长,简单或者有分枝,直,锯齿状、波曲状、顶端有钩或者卷曲。某些种相邻的分生孢子梗常常发生菌丝融合。许多情况下,瓶梗和产孢的分枝可以从平展的产孢区中未分化的菌丝上产生,产孢细胞瓶形,通常在分生孢子梗的顶端呈二歧式轮状排列,较少单生;或者在分生孢子梗和分枝隔膜下面呈涡状排列,分枝柱形、钻形、烧瓶形、坛形或者亚球形,在分生孢子下面常常是细的短柱形颈。总起来说,木霉的分生孢子梗按照分枝形式可以分为4种类型,即粘帚霉型( Gliocladium-like)、厚基孢型( Pachybasium-like)、轮枝菌型( Verticillium-like)和木霉型( Trichoderma-like),分生孢子梗的分枝形式是鉴定木霉的重要形态指标。
分生孢子单细胞,无色,浅灰色,绿色或者棕色,壁光滑或有明显粗糙,某些种在外壁上有波状、疱状或者翅形突起饰物,亚球形、卵形、椭球形、长椭球形或者短柱形;或者聚集为球形的头状体,或者聚集在囊状的包被中(两个种有这种结构)。如果出现有性阶段,则为 Hypocrea。
以上描述主要依据Bissett于1984和1991年发表的研究论文,也参考了Chaverri等(2004)的相关描述。相关的培养特征、产孢簇形态以及分生孢子梗的形态特征见有关图片。
模式种: Trichoderma viride Pers.:Fr.。
2. 木霉属的概念
木霉属( Trichoderma)的概念最早由Persoon(1794)引入,用于描述4个在宏观上特征相似的真菌,Persoon描述的这4个真菌为由毛发状覆盖物包裹的粉状结构,最先描述的 Trichoderma有4个种,即 T. aureum,T. nigrescens,T. roseum和 T. viride(具有不同的分生孢子颜色),现在已知它们是彼此无关的种类。 Trichoderma这个名字现在仅仅用于描述以 T. viride为代表的分布广泛的绿色类型。依据微观特征,Rifai(1969)给予 Trichoderma属的界定,现在已经被普遍接受。木霉属( Trichoderma)的分生孢子梗为重复多次分枝,常常排列成规则的树状结构,即在多级水平上重复地无限分枝,初级分枝产生一些小的次级分枝,次级分枝也可以再次分枝,如此重复不已。最早形成的分枝较长,连续形成的分枝向顶以及随着离基部距离的增加而逐步变短,所有水平的分枝和瓶梗均为二歧式。瓶梗为安瓿形至坛形,基部常常缢缩,在中间部位多少有些膨大,在近顶端突然变细而形成近柱形的颈部。瓶梗的排列为轮状,位于分生孢子梗分枝的终端,有时呈单生,或者呈涡状排列,位于靠近分生孢子梗与其分枝分隔的下方。 Trichoderma的分离物可以依据其宏观特点进行辨别,即在培养基上生长迅速,稀疏的气生菌丝,以及典型的白色或者绿色的产孢堆。
已知的 Trichoderma有性型为 Hypocrea,在 Podostroma中也可能存在,以及与 Hypocrea相近的一些属中。Doi曾经观察了 Hypocrea以及与 Hypocrea有关的60 多个种。许多 Hypocrea种类产生典型的 Trichoderma无性型(在人工培养时),其他一些 Hypocrea种则产生具有更为不规则的分生孢子梗分枝方式的无性型,这些无性型则不易认定为 Trichoderma,而是收到 GliocladiumCorda、 VerticilliumNees或 AeremoniumLink:Fr.。
多数土壤以及木材上生长的 Trichoderma菌株的有性型未知,很可能 Trichoderma中的许多常见种类没有有性世代。Domasch(1980)认为(在一篇文献综述中)将 Trichoderma无性型与单一的有性型联系起来,通常是不可能的。Rifai的每个集合种可以与不同的有性世代种相关联,而这些有性世代种的不完全阶段在人工培养时难以区别。他们还断言,不能保证将来会把 Trichoderma进一步划分。
Rifai(1969)根据瓶梗以及分枝产生的角度将 Trichoderma和 Gliocladium区别开。 Gliocldium的分枝为帚状,或者瓶梗紧密地贴在一起形成聚集的头状体, Gliocladium的模式种是 G. penicillioides Corda,具有典型的帚状分枝的分生孢子梗,分枝局限在分生孢子梗的顶端(Corda,1984)。相对于 Trichoderma 来说, Gliocladium的瓶梗更为细长,聚集更为紧密。已知的 Gliocladium的有性世代属于 SphaerostilbellaP. Hennings以及 Hypocreales的有关属(Seifert,1985)。 Hypocrea pallida Ell. & Ev. 以及与 Gliocladium有关的无性型被Samuels & Seifert(1990)归到 Sphaerostibella,Gliocladium roseum(Link)Bainier(有性世代为 Nectria ochroleuca(Schwein)Berk)和其他在形态上相似的种类之区别在于分生孢子形成覆瓦状的柱形结构,而不是胶质的头状结构。 ClonostachysCorda(1839)可能包括这些种类,它们的有性型是 Nectria(Samuels,1988)。
Rifai(1969)将 Hypocrea gelatinosa(Tode:Fr.)Fr.的无性型归类于 Gliocladium,同 T. virens一样。在这两个种中,分枝和瓶梗有时聚集,相邻瓶梗上产生的分生孢子可以联结为一个大型的分生孢子球,但是, T. virens和 H. gelatinosa的粗大分生孢子梗有重复多次的树状无限分枝,而不是帚状的二歧式分枝或者瓶梗。后面两个种的分生孢子梗分枝方式和膨大的瓶梗比起 Gliocladium更像 Trichoderma,因此这两个种也归在 Trichoderma 里。
Rifai(1969)还从 Trichoderma中除去了 Hypocrea lactea(Fr:Fr)Fr.以及 H. hanuaDingley的无性型。他将这两个种归在 VerticilliumNees中,比起 Trichoderma,该属的瓶梗更长,而且排列为规则的轮状。在 VerticilliumS. str中,从粗大分生孢子梗的主轴上产生的初级分枝通常仅有一次二级分枝,终端以轮状排列的钻形瓶梗结束。Hughes(1951)描述了其模式种 Verticillium luteo-album(Link:Fr.)Subram、 Hypocrea,包括 H. laetea和 H. nunua的无性型,可以依据分生孢子梗的分之形式与 Verticillium相区别,在 Hypocrea的无性型中,分枝方式为树状,初级分枝产生二级小分枝,而小分枝可以再次分枝。因此,现在的处理是, H. laetea和 H. hunua包括在 Trichoderma中,虽然其分生孢子梗不像传统上认为的 Trichoderma那样具有复杂的规则分枝。
在 Hypocrea无性型的形态学方面有渐变现象,有些种类产生不规则的分生孢子梗,分生孢子梗产生相对较长的瓶梗,而有些种类则有规则的树状分枝以及具有明显膨大的瓶梗(更像典型的 Trichoderma)。 H. laetea和 H. hunuaDingley的分生孢子梗具有较少不规则的分枝,分枝的瓶梗的排列可以呈帚状,或者单生,间距也不规则。分枝可以是二歧式的,但是有些时候分枝和瓶梗聚集在一起,或者相互间平行排列。瓶梗类似钻状,与典型的 Trichoderma相比,瓶梗更长,虽然有一些是明显膨大的类型。在 H. atrogelatinosaDingley中,其瓶梗比 H. laetea和 H. hunua短,而且分之和瓶梗通常对生或者为帚状排列。 H. vinosaCook的瓶梗很多是膨大型的,而且排列为二歧式的轮状。 H. vinosa的一些菌株具有规则树状分枝的分生孢子梗,其上着生近似安瓿形的瓶梗,在终端以二歧式轮状排列。 H. rufa的分生孢子梗则呈规则的树状分枝,其分枝和瓶梗聚集,瓶梗的形状一般为安瓿形。
如果考虑 Hypocrea无性型的所有形态学变异,那么,要把以 T. viride 为模式种的 Trichoderma与其他连续渐变的类型相区别是很困难的。最后,在本研究中, Hypocrea的所有无性型都包括在了 Trichoderma中。 Trichoderma和 Hypocrea无性型的总体变异幅度是很大的,但是,利用分生孢子梗具有二歧式树状分枝这个特点,可以把这些种类与 Gliocladium和 Clonostachys(具有帚状分枝的分生孢子梗)以及 Verticillium(具有轮状或者二歧式轮状分枝)区别开。
Bissett (1984,1991a,1991b,1991c,1992;Gams &Bissett,1998)修订了Rifai的集合种分类方法,主要依据是形态特征的细致判断,将形态学标准扩展到了更大的范围,包括了某些Hypocra种类无性型所具有的形态学变化,同时还吸纳了以前鉴定 Gliocladium时采用的一些形态特征;还建立了五个属下分类单元(Bissett,1991a)即组: Longibrachiatum、Pachybasium、Trichoderma、Saturnisporum和 Hypocreanum。分子工具在真菌分类中的出现,促使研究人员在上世纪90年代中期重新审视基于形态学的木霉分类系统。G.J. Samuels (Beltsville,MD,USA)、T. Börner (Berlin,FRG)和C.P. Kubicek (Vienna,Austria)实验室合作,率先对Bissett的 Longibrachiatum组进行了修订,他们使用的技术包括多种分子标记技术(ITS1和ITS2序列分析,RAPD),生理学(同工酶分析)和表型特征,并且首次包括了该组 Trichoderma种类可能的有性型作为研究对象(Kuhls et al .,1996;1997;Samuels,1996;Samuels et al .,1998;Turner et al .,1997)。结果是, Longibrachiatum组在系统发育上是独立的,并且包括10个分类单元,其中有4对为无性-有性关系: H. schweinitzii/ T. citrinoviride、 H. pseudokoningii/ T. pseudokoningii、H. jecorina/ T. reesei和 H. orientalis/ T. longibrachiatum。还进一步将 Saturnisporum组(只包括两个种即 T. saturnisporum和 T. ghanense (Doi et al .,1987))合并在 Longibrachiatum组中,确认 T. ghanense为 T. parceramosum的同义名。但是,总体上来说Bissett (1984)所建立的 Longibrachiatum组的概念得到确认,表明木霉的形态分类和分子分类技术之间具有密切关系。 Longibrachiatum组是木霉中较小的组,系统发育上与其他组的关系比较远。当研究木霉中较大的组时,形态特征与分子系统发育之间的关系就变的复杂。Kindermann et al .(1998)第一次试图分析整个木霉属的系统发育,采用的方法是对rDNA的ITS1区进行序列分析,发现 Pachybasium组实际上是并系的(paraphyletic)组群或者分类单元。这一结果被后来用多个基因进行的研究结果所证实(Kullnig-Gradinger et al .,2002;Chaverri et al .,2003b)。值得注意的是,按照这样的结果,所命名的 Pachybasium组模式菌株 Pachybasium hamatum(Bonord.)= T. hamatum不再是两个 Pachybasium进化枝中主要分枝(clade B)的成员(Kindermann et al .,1998),在一个进化枝中的簇加上 T. pubescens和 T. strigosum几乎包括了 Trichoderma组所有的种类( T. aureoviride除外)。由于 Pachybasium的A和B进化枝之间形态学上缺乏明显的界限(Kindermann et al .,1998),现在还用 Pachybasium来命名这两个进化枝。除了上述三个种外, Pachybasium B包括了Bissett (1991b)归类到该组的所有分类单元。另外, Pachybasium B的问题是有些种类遗传变异非常明显,例如 T. harzianum。由于在田间发现的该组种类占据了大多数(Kullnig et al .,2000;Kubicek et al .,2003;Wuczskowski et al .,2003;Gherbawy et al .,2004;Druzhinina et al .,2004b),因此如何鉴定该组种类就成了一个重要问题。
总起来说,木霉( Trichoderma)具有透明的瓶梗,产生于外露的可育菌丝分枝或者分生孢子梗上,分生孢子一般光滑,很少有饰物,典型形状为椭圆形至近似长方形,很少为球形,大多数为绿色或者透明,很少为黄色。如果产生厚垣孢子,其典型形状为球形至亚球形,产生于菌丝中间或者菌丝尖端。在 T. stromaticum Samuels中,后垣孢子则包含细胞球,也能形成典型的厚垣孢子(Samuels et al .,2000)。 Trichoderma virens (Miller et al .) Arx 的部分特征是在纯培养条件下形成大量的厚垣孢子(Chaverri et al .,2001a)。
3.木霉属的有性阶段
木霉的有性阶段一般发现为肉座菌属( Hypocrea),Tulasne于1865年即提出了肉座菌属与木霉属的关系,之后Tulasne于1865年,Brefeld于1891年证实了绿色木霉是红棕肉座菌( H. rufa)的分生孢子阶段。Bisby于1939年报道 H. rufa和胶质肉座菌( H. gelatinosa)具有无法区别的木霉型分生孢子梗,而 H. pulvinata则具有头孢霉型( Cephalosporium-type)的分生孢子梗。
肉座菌属属于子囊菌亚门(Ascomycotina)的核菌纲(Pyrenomycetes)、球壳目(Sphaeriales)、肉座菌科(Hypocreaceae),腐生于树皮、木材和高等真菌的老龄子实体上。该属由Fries于1825年建立,子座肉质或者木栓质,平展伏生,多数为垫状、盘状或者半球状,有柄或者无柄,颜色鲜亮,有的也呈暗褐色;子囊壳完全埋没在子座内,很少为半裸露状态,子囊壳孔口基本不突出于子座外,每一个子囊有8个双细胞的孢子,即每一个孢子可以分为两个圆形细胞,称为分(子囊)孢子,因此成熟的子囊含有16个孢子。每对分孢子中远基的称为远基分孢子,近基的则称近基孢子。分孢子常为圆柱形、倒卵形、球形或者椭圆形,透明无色或者淡绿至深绿色,表面粗糙或者具有瘤状突起。
19世纪中期,已经发现了木霉与肉座菌( Hypocrea spp.)之间的关联(Tulasne,LR.,Tulasne,C. 1865),Doi et al.(1979)提出已知木霉的有性阶段与子囊菌纲 Hypocrea, PodostromaKarst和 Sarawakus密切相关,Rehner & Samuels (1995)以及其他学者(Chaverri,Castlebury,Overton,Samuels,2004; Kullnig-Gradinger,Szakacs,& Kubicek,2002)则发现,有些木霉的种类,以及其他尚未发现相关有性型的非木霉属的某些成员,仅依据其有性阶段难以相互区别。后来通过更多的野外观察工作逐步发现,很多木霉的种类与肉座菌有关,肉座菌为木霉的有性型,例如 T. harzianum以及 H. lixii的无性型(Chaverri & Samuels,2003), T. virens, H. virens的无性型等(Chaverri,Samuels & Stewart,2001)。但是,一些分布广泛并且重要的种类,例如 T. asperellum,至今还没有发现有关的有性型,因此属于无性繁殖的类型(Samuels,2006)。从目前的研究资料来看,多数木霉种类的有性阶段属于肉座菌属真菌。由于木霉菌的分生孢子在人工培养条件下很难形成有性阶段的子实体,木霉菌研究专家运用包括等位酶技术、RAPD、UP-PCR、DNA指纹和序列分析技术在内的分子生物学方法,通过比较木霉菌与其有性型真菌(主要是肉座菌属)DNA 序列的相似性程度,来判断两者的无性型与有性型的关系(Samuels et al.,1994,Leuchtmann et al.,1996;Zimand et al.,1994;Lübeck et al.,1999;Meyer et al.,1992);从肉座菌属的子囊孢子开始培养,虽然形成了类似木霉菌的菌落,但很多菌株难以鉴定到木霉属内的种(Dodd et al.,2002)。
Hypocrea 于1825年由Elias Fries首先描述,其描述基础是产生透明子囊孢子的 Sphaeria rufa Pers.:Fr.。现在,该属的模式种位 Hypocrea rufa (Pers. :Fr.)Fr.。 Hypocrea 的主要特征是子囊壳包埋在肉质的子座中,子座由拟薄壁组织或者高度紧密排列的菌丝组成。 Hypocrea 发育早期具有8个具有1个分隔的子囊孢子,然后在隔膜处两个细胞相互脱离,产生16个分子囊孢子。虽然从隔膜处发生断裂现象是该属的显著特点之一,但是其他属的真菌例如 Aphysiostroma、Arachnocrea、Dialhypocrea、Podostroma、 Protocrea、 Pseudohypocrea以及 Sporophagomyces,也具有断裂产生的子囊孢子(Rossman et al.,1999)。新近的DNA序列研究表明, Podostroma以及 Protocrea与 Hypocrea同系。 Podostroma包括的种类具有直立的子座柄。一些研究发现 Podostroma与 Hypocrea之间具有相似性,其主要差别在于前者的子座具有柄(Boedijn,1934;Doi,1966;Rossman et al .,1999)。 Podostroma其他方面的特点,例如子座组织,子囊壳以及无性型的形态,还有生态特点等,与 Hypocrea没有明显区别。 Aphysiostroma、Arachnocrea、Dialhypocrea、Pseudohypocrea以及 Sporophagomyces等,与 Hypocrea相似,都具有断裂的子囊孢子,有性型和无性型的形态也较独特,其中最为显著的现象是, Arachnocrea,Pseudohypocrea,和 Sporophagomyces的子囊孢子为双锥形。 Arachnocrea和 Sporophagomyces则为真菌寄生类型。最近的分子系统发育研究表明, Aphysiostroma和 Arachnocrea位于 Hypocreas. str.的基部(Põldmaa et al .,1999,Zhang & Blackwell,2002)。 Hypocrea pallida Ellis & Everh是广义 Aphyllophorales类群的寄生物,由于具有2个细胞的子囊孢子,以及子囊壳包埋在假子座或菌丝层中,从而被置于 Hypocrea属。但是, H. pallida的无性型为粘帚霉类型,与 G. penicillioides Corda相似,后者为 Gliocladium的模式种,也与 Sphaerostilbella aureonitens (Tul.)Seifert et al .相似。分子研究表明,相对于 Hypocrea来说, H. pallida与 Hypomyces以及 Sphaerostilbella的关系更加密切(Põldmaa et al .,1999)。
子囊孢子的颜色也用来描述 Hypocreales科内属的特征,其中 Creopus Link和 Chromocrea Seaver两个属,由于具有绿色的子囊孢子而独立于 Hypocrea。在1791年,将 Sphaeria gelatinosa Tode描述为 H. gelatinosa (Tode:Fr.)Fr. Link (1833),该种具有绿色的子囊孢子,曾经被Fries (1849)放在 Hypocrea属内,Tode的依据是他自己关于 S. gelatinosa上 Creopus属的描述。虽然Link没有明确指出 Creopus的独立基于其绿色的子囊孢子,但是后来放在该属里面的种类都具有这一特征。 Creopus属共有4个种,即 Cr. gelatinosus (Tode:Fr.) Link 1833、 Cr. palmicola (Berk. & Cooke) Boedjin 1951、 Cr. spinulosus (Fuckel) Moravec 1956和 Cr. velenovskyi Z. Moravec 1956。后来,Seaver (1910)建议用 Chromocrea来包括具有绿色子囊孢子的 Hypocrea种类,他指定 C. gelatinosa为模式种,因此 Chromocrea就成为 Creopus后来的一个同模标本的同物异名。该属具有8个种,即 Ch. aureoviridis(Plowr. & Cooke ) Petch 1938, Ch. Ceramica (Ellis & Everh.) Seaver 1910, Ch. cupularis (Fr.) Petch 1938, Ch. gelatinosa(Tode:Fr.)Seaver 1910, Ch. marathwadi Tilak & Gaikwad 1978, Ch. Nigricans Imai 1935, Ch. Spinulosa (Fuckel) Petch 1950和 Ch. Substipitata Seaver 1910。 Sarawakus 是唯一的另外一个具有绿色子囊孢子,形态上类似于 Hypocrea的属。在 Hypocreales中,除了 Hypocrea外,只有 Viridispora Samuels & Rossman ( Nectriaceae)的成员以及 Neonectria的一些成员具有绿色的子囊孢子。有关 Hypocrea的第一个“现代化”技术处理结果,是Dingley (1952,1957)发表的,她描述了子座的解剖学,也进行了人工培养观察,并将种与其无性型相互对接关联。在新西兰,描述了 Hypocrea的许多新种及其无性型(Dingley,1952、1957)。Dingley将所有 Hypocrea的无性型鉴定为典型的 T. viride Pers. :Fr.,虽然在其图片中可以见到彼此之间具有明显的不同。在Webster (1964),Webster & Rifai (1968)以及Rifai & Webster (1966a,b)工作的基础上,Rifai (1969)发表了有关 Trichoderma的重新检查后的分类结果,在木霉的分类研究工作中,这是十分重要的里程碑式研究成果,他将木霉分为了9个集合种‘aggregate species’。此后,Doi在日本出版了 Hypocrea及其相关种类的专论(Doi,1969、1972),这是迄今为止有关 Hypocrea研究的最大型的研究工作。在该专论中, Hypocrea的种类都有描述和图示。Doi及其同事基于在日本、新几内亚和南美洲收集的标本,描述了许多 Hypocrea的新种以及他们的无性型。(Doi,1966、1968、1969、1971、1972、1973、1975、1976、1978;Doi & Doi,1980;Doi & Yamatoya,1989)。
以子座的解剖学特点为主要依据,参考无性型以及子囊孢子的着色情况,Doi建议了一个 Hypocrea的属下分类系统。依据有性型的特点,Doi描述了两个亚属,即 H.subg. Heterocrea和 H.subg. Hypocrea。在 H.subg. Hypocrea中,又定义了两个组,即 H. sect. Homalocrea和 H.sect. Hypocrea。Doi (1972)又确立了亚组 H. subsect. Creopus,属于亚属 H.sect. Hypocrea,并把大多数 Hypocrea具有绿色子囊孢子的种包括在其中。在这个亚组中,他又将种区别为3个“人工组”,即Aureoviridis、Cornea和Gelatinosa,其分组依据是子座类型和分生孢子梗的分枝类型。Aureoviridis组的种类,其子座脆而易碎,表面一般没有突出物,无性型为木霉( Trichoderma)或者厚基孢( Pachybasium)类型(Bissett,1991a、b)。Cornea组的特征是其子座为革质,无性型为近似轮枝菌( Verticillium)类型。Gelatinosa组的特点是子座脆而且易碎,一般具有表面突出物,无性型为 Gliocladium类型。近期的传统和分子研究结果显示,具有 Trichoderma、 Pachybasium、 Gliocladium或者 Verticillium类型的无性型的 Hypocrea种类,并非单系组群(Kindermann et al .,1998;Kullnig-Gradinger et al .,2002)。有性型表型类型的进化趋同现象十分明显,因此,系统发育研究不支持Doi有关 Hypocrea的属下分类系统(Chaverri et al .,2000;Kullnig-Gradinger et al .,2002;Lieckfeldt et al .,1998a)。
大多数 Hypocrea种类是在19世纪和20世纪早期描述的,当时所依据的标本大多来自旅行家、传教士、病理学家或者其他野外工作者,标本一般送给相关专家进行鉴定描述,例如英国的M.J. Berkeley、德国的P. Hennings和美国的J.B. Ellis等专家。这种现象是由当时的时代特点所决定的,描述的新种一般发表于包含了大量不同真菌的大型著作中,而且这些描述都没有匹配图示,只有简单的描述性文字以及有关基物的注解。比较规范的描述包括了子囊孢子的测量结果,但都没有将无性型与标本相关联的记载,或者对子座解剖学的详细描述。Dingley首先在系统学研究中使用了有性型解剖学特点,Doi后来也采用了这些特征,这比以前的简单化描述前进了一大步。总而言之,有性型是子囊菌分类系统所依据的基础,而且是全型(即有性型+无性型)中的名称的载体部分(International Code of Botanical Nomenclature,条款59)。但是,随着子囊菌中有性型与无性型关联研究资料的丰富(包括肉座菌与木霉),以及肉座菌系统发育研究中分子技术的引入,越来越多的研究结果显示,肉座菌的子座具有高度的保守结构,在种水平上的分类中用处不大,不能作为细致的分类与鉴定依据。例如,Samuels et al . (1998)在其有关 Hypocrea schweinitzii (Fr.)Sacc. 集合体的专论中,表明 H. schweinitzii的广义有性型之间几乎无法区别。种类之间虽然差别明显可见,但这些差别是在无性型、生长特性以及DNA序列方面,而不是在有性型的形态方面。在包括相近种类研究的例子中,不同种类之间的显著区别在木霉无性型阶段明显可见,但在有性型形态和解剖学特征方面就不那么明显。
总结最近的研究结果,得出如表4-1所示的木霉-肉座菌各种类之间的对应关系,它们在进化枝中的位置也一并列出(Druzhinina & Kubicek,2005)。可以看出,许多木霉种类尚未发现所对应的有性型,而有的有性型,也没有对应的木霉种类。肉座菌的无性型除了木霉类型的分生孢子梗之外,还有 Cephalosporium、Verticillium、Gliocladium和 Pachybasium的分生孢子梗类型。
分布种类


