视杆细胞(位于眼球视网膜内的细胞)

VLoG
次浏览
更新时间:2023-05-22
视杆细胞
位于眼球视网膜内的细胞
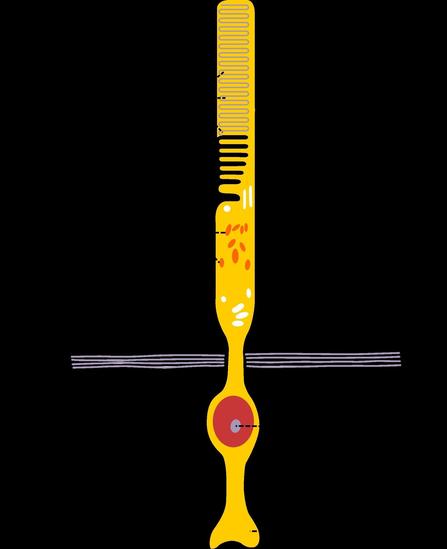
视杆细胞是位于眼球视网膜内的细胞,其作用是分辨黑白,动态信息。
基本信息
| 中文名 | 视杆细胞 |
| 外文名 | rod cell |
| 类型 | 视觉细胞 |
| 作用 | 分辨黑白,动态信息 |
简介
视杆细胞
,称为视杆,视杆外节的膜盘除基部少数膜盘仍与胞膜相连,其余大部分均在边缘处与胞膜脱离,成为独立的膜盘。膜盘的更新是由外节基部不断产生,其顶端不断被色素上皮细胞所吞噬。膜盘上镶嵌有感光物质,称视紫红质(rhodopsin),能感受弱光。视紫红质是由11-顺视黄醛(11-cisretinae)和视蛋白(oposin)组成,前者是维生素A的衍生物,当维生素A缺乏时,视紫红质合成不足,则患夜盲症。视杆细胞体较小,核圆形染色较深,其轴突末梢不分之呈球型,与双极细胞的树突形成突触。
约翰·霍普金斯大学科学家领导的研究小组发现,眼睛感光的任务极有可能仅由三种细胞负责。
在2003年6月15日期《自然》杂志的网络版上,研究小组报道,视锥细胞、视杆细胞和产生黑视蛋白的特殊视网膜细胞一起合作,包揽了小鼠对光强做出反应的全部工作。有其他研究人员提出产生感光色素cryptochrome的细胞也有感光作用,但霍普金斯的科学家表示,就小鼠而言,cryptochrome细胞没有这个作用,在人类中恐怕也是如此。
“我们相当确信,视锥/视杆细胞系统和黑视蛋白系统是哺乳动物眼部唯一两个感光系统。”约翰·霍普金斯大学基础生物医学科学研究所的神经科学教授King-Wai Yau博士说。“永远不要说永远,但至少目前还没有证据表明有第三个系统存在。”
视紫红质分解
在实验中,Yau和博士后研究人员Samer Hattar博士除去编码三种关键蛋白的基因,三种蛋白中的每一种都有助于传递来自视杆细胞、视锥细胞和黑视蛋白细胞的光信息。早先,阻止小鼠模型视杆细胞、视锥细胞和产生视力的细胞的光信息传递的传统方法是使视杆细胞和视锥细胞退化。
“但我们想避免传统的视杆、视锥细胞退化孝小鼠模型所带来的不确定性。”Hattar说。“在那个模型中,你无法确定视杆细胞和视锥细胞丧失的百分比,也无法确定退化的视网膜本身是否会影响你的观察结果。”
与伦敦帝国学院的Robert Lucas合作,霍普金斯大学的研究人员证明,3基因敲除小鼠根本无法调节瞳孔对光做出反应。在多伦多大学进行的实验中,文章合着者Nicholas Mrosovsky发现,3基因敲除小鼠暴露于光照中时,活跃水平的调节也不适当。而三种蛋白各含一半剂量的小鼠在两个实验中反应都正常。
“即使其内部生物钟告诉它现在是夜晚,正常小鼠暴露于强光时也会表现得不太活跃,会藏起来甚至睡觉。”Yau说。“这被看作是动机发生改变,也许小鼠意识到在光照中--即使生物钟告诉它们现在是夜晚,它们被捕食的机会更大。但3基因敲除小鼠好像在白天与夜晚一样活跃。”
研究人员设计了另外一个实验来排除cryptochrome蛋白有感光作用,据报道该蛋白与果蝇的感光有关。由于每个光敏蛋白都是对光的特征波长最敏感,因而将小鼠暴露于单波长光中可以揭示对眼睛的非视觉功能最重要的蛋白。
在这些在伦敦帝国学院进行的实验中,研究人员发现传统的视杆/视锥细胞退化小鼠当暴露于黑视蛋白细胞而不是cryptochrome细胞感应的光时,“重设”内部生物钟的能力最强。
“在果蝇中,有两个以上系统负责感光,而在斑马鱼和鸟类等一些动物中,感光甚至不只局限于眼睛,而这对于传统的视杆/视锥细胞退化小鼠而言是不可能的事情。”Hattar说。Hattar正在区分视杆细胞和视锥细胞在感光中所扮演的角色。“但在哺乳动物中,所有这些功能--视力、生物钟和光诱导活动,都存在于一个部位,那就是视网膜,并且仅由两个系统、三类细胞完成。”
盲人的视杆细胞和视锥细胞丧失功能,但眼睛中产生黑视蛋白的细胞似乎仍能提供足够的信息使身体和大脑与生物钟协调一致。但如果完全失去双眼,生物钟就会遭到破坏。
视觉形成过程
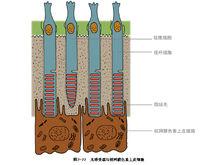
哺乳动物光感受器细胞模式图光感受器按其形状可分为两大类,即视杆细胞和视锥细胞。夜间活动的动物(如鼠)视网膜的光感受器以视杆细胞为主,而昼间活动的动物(如鸡、松鼠等)则以视锥细胞为主。但大多数脊椎动物(包括人)则两者兼而有之。视杆细胞在光线较暗时活动,有较高的光敏度,但不能作精细的空间分辨,且不参与色觉。在较明亮的环境中以视锥细胞为主,它能提供色觉以及精细视觉。这是视觉二元理论的核心。在人的视网膜中,视锥细胞约有600~800万个,视杆细胞总数达1亿以上。它们似以镶嵌的形式分布在视网膜中;其分布是不均匀的,在视网膜黄斑部位的中央凹区,几乎只有视锥细胞。这一区域有很高的空间分辨能力(视锐度,也叫视力)。它还有良好的色觉,对于视觉最为重要。中央凹以外区域,两种细胞兼有,离中央凹越远视杆细胞越多,视锥细胞则越少。在视神经离开视网膜的部位(乳头),由于没有任何光感受器,便形成盲点。
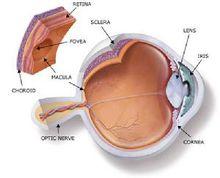
视网膜内有感光细胞层,人类和大多数脊椎动物的感光细胞有视杆细胞和视锥细胞两种。感光细胞可通过终足和双极细胞发生突触联系,双极细胞再和神经节细胞联系,由节细胞发生的突起在视网膜表面聚合成束,然后穿过脉络膜和巩膜后构成视神经,视神经出眼球后穿视神经管入颅腔,经视交叉连于间脑。
目前认为,物像落在视网膜上首先引起光化学反应,已从视网膜上提取出感光物质。这些物质在暗处呈紫红色,受到光照时则迅速退色而转变为白色。如将蛙或兔放在暗室中,使动物眼朝向明亮的窗子一定时间,然后遮光立即摘出眼球,剔出视网膜,用适当化学物质如明矾处理视网膜,则可发现动物视网膜留有窗子的图像,窗子的透光部分呈白色,窗框部分呈暗红色。这些都说明视网膜上感光物质在光线作用下所出现的光化学反应。在感光细胞的大量研究中,对视杆细胞研究得比较清楚。视杆细胞的感光物质称为视紫红质,它由视蛋白和视黄醛结合而成。视黄醛由维生素A转变而来。视紫红质在光照时迅速分解为视蛋白和视黄醛,与此同时,可看到视杆细胞出现感受器电位,再引起其他视网膜细胞的活动。
视紫红质在亮处分解,在暗处又可重新合成。人在暗处视物时,实际上既有视紫红质的分解,又有它的合成。光线愈暗,合成过程愈超过分解过程,这是人在暗处能不断看到物质的基础。相反在强光作用下,视紫红质分解增强,合成减少,视网膜中视紫红质大为减少,因而对弱光的敏感度降低。故视杆细胞对弱光敏感,与黄昏暗视觉有关。视紫红质在分解和再合成过程中,有一部分视黄醛将被消耗,主要靠血液中的维生素A补充。如维生素A缺乏,则将影响人在暗处的视力称为夜盲症。
结构
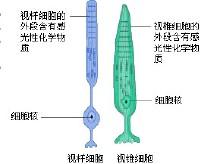
视杆细胞和视锥细胞
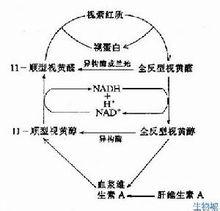
视杆细胞的视色素叫做视紫红质,它具有一定的光谱吸收特性,在暗中呈粉红色,每个视杆细胞外段包含109个视紫红质分子,视紫红质是一种色蛋白,由两部分组成。其一是视蛋白,有348个氨基酸,分子量约为38000;另一部分为生色基团——视黄醛,是维生素A的醛类,因为存在若干碳的双键,它具有几种不同的空间构型。在暗处呈扭曲形的11-型异构体,但受光照后即转变为直线形的全-反型异构体。后者不再能和视蛋白相结合,经过一系列不稳定的中间产物后,视黄醛与视蛋白相分离。在这一过程中,视色素分子失去其颜色(漂白)。暗处它在酶的作用下,视黄醛又变为11-顺型,并重新与视蛋白相结合(复生),完成视觉循环。在强光照射后,视紫红质大部分被漂白,其重新合成需要约1小时。随着视紫红质的复生,视网膜的对光敏感度逐渐恢复,这是暗适应的光化学基础。当动物缺乏维生素A时,视觉循环受阻,会导致夜盲。
信息处理
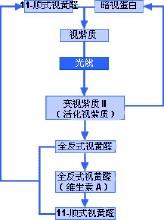
视紫质化学反应图解
视杆细胞的信号和视锥细胞的信号,在视网膜中的传递通路是相对独立的,直到神经节细胞才汇合起来。接收视杆细胞信号的双极细胞只有一类(杆双极细胞),但接收视锥细胞信号的双极细胞,按其突触的特征可分为陷入型和扁平型两种,这两种细胞具有不同的功能特性。在外网状层,水平细胞在广阔的范围内从光感受器接收信号,并在突触处与双极细胞发生相互作用。此外,水平细胞还以向光感受器反馈的形式调制信号。在内网状层双极细胞的信号传向神经节细胞,而无长突细胞则把邻近的双极细胞联系起来。视杆和视锥细胞信号的汇合也可能发生在无长突细胞。
感光原理
感光特点
光线进入眼睛后,首先通过角膜,然后分别透过房水、晶状体和玻璃体。最后,光线到达眼睛的感光组织视网膜。视网膜中有两种细胞分别叫做视杆细胞和视锥细胞。视杆细胞负责昏暗光线下的视物,而视锥细胞则负责处理色彩和细节。当光线接触到这两种细胞时,会发生一系列复杂的化学反应。形成的化学物质(活化视紫质)会在视神经中产生电子脉冲。一般来说,视杆细胞的外段细长,而视锥细胞的外段更像是锥形。
感光物质
这两种细胞的外段都含有感光性化学物质。在视杆细胞中,这种化学物质叫做视紫质;而视锥细胞中的则叫做色素。光线进入眼睛后,会接触到感光性化学物质视紫质(也叫做视紫红质)。视紫质是暗视蛋白和11-顺式视黄醛的混合物,11-顺式视黄醛来源于维生素A。因此,缺乏维生素A就会导致视力问题。视紫质遇光会发生分解,因为光会导致视紫质中的11-顺式视黄醛发生物理变化,转化为全反式视黄醛。第一个反应只需一万亿分之几秒的时间。11-顺式视黄醛是角状分子,而全反式视黄醛是直线分子。因此,化学性质极不稳定。视紫质可分解为几种中间化合物,但最终(在不到一秒的时间里)会形成变视紫质II(活化视紫质)。这种化学物质能够产生电子脉冲,电子脉冲传输到大脑后,大脑将其解译为光线。

视杆细胞
1.视杆细胞的细胞膜(外层)带有电荷。当光线激活视紫质时,它就会导致环式GMP(鸟苷单磷酸)减少,而电荷随之增加。这样就会在细胞周围产生电流。随着光线的增多,就会有更多的视紫质被激活,也就会产生更强的电流。
2.最后,电子脉冲被传到神经节细胞,再传到视神经。
3.这些视神经在视交叉处汇集。在这里,视网膜内部的神经纤维会通往另一侧大脑,而视网膜外部的神经纤维则通往同侧大脑。
最后,必须重新形成视紫质,才能重复进行产生视觉的过程。全反式视黄醛转化成11-顺式视黄醛,然后与暗视蛋白结合后,便形成视紫质,视紫质遇光后便会重新开始该过程。
感光换能机制
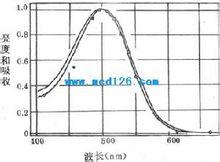
弱光条件下人眼所感到的光谱亮度曲线
(一)视紫红质的光化学反应及其代谢
弱光条件下人眼所感到的光谱亮度曲线视紫红质的分子量约为27-28kd,是一种与结合蛋白质,由一分子称为视蛋白(opsin)的蛋白质和一分子称为视黄醛(retnal)的生色基团所组成。视蛋白的肽链序列已搞清,它的肽链中有7段穿越所在膜结构、主要由疏水性氨基酸组成的α-螺旋区段,同一般的细胞膜受体具有类似的结构。视黄醛由维生素A变来,后者是一种不饱和醇,在体内一种酶的作用下可氧化成视黄醛。提纯的视紫红质在溶液中对500nm波长的光线吸收能力最强,这与人眼在弱光条件下对光就业上蓝绿光区域(相当于500nm波长附近)感觉最明亮(不是感到了蓝绿色)的事实相一致,说明人在暗视觉与视杆细胞中所含视紫红质的光化学反应有直接的关系。
视紫红质在光照时迅速分解为视蛋白和视黄醛,这是一个多阶段的反应。目前认为,分解的出现首先是由于视黄醛分子在光照时发生了分子构象的改变,即它在视紫红质分子中本来呈11-顺型(一种较为弯曲的构象),但在光照时变为全反型(一种较为直的分子构象)。视黄醛分子构象的这种改变,将导致视蛋白分子构象也发生改变,经过较复杂的信号传递系统的活动,诱发视杆细胞出现感受器电位。据计算,一个光量子被视紫红质吸收,就足以使视黄醛分子结构发生改变,导致视紫红质最后分解为视蛋白和视黄醛。视紫红质分解的某些阶段伴有能量的释放,但这看来不是诱发感受器电位的直接原因。
在亮处分解的视紫红质,在暗处又可重新合成,亦即它是一个可逆反应,其反应的平衡点决定于光照的强度。视紫红质再合成的第一步,是全反型的视黄醛变为11-顺型的视黄醛,很快再同视蛋白结合。此外,贮存在视网膜的色素细胞层中的维生素A也是全反型的,它们也可在耗能的情况下变成11-顺型的,进入视杆细胞,然后再氧化成11-顺型的视黄醛,参与视紫红质的合成补充;但这个过程进行的速度较慢,不是促进视紫红制裁再合成的即时因素。人在暗处视物时,实际是既有视紫红质的分解,又有它的合成,这是人在暗光处能不断视物的基础;光线愈暗,全盛过程愈超过分解过程,视网膜中处于合成状态的视紫红质数量也愈高,这也使视网膜对弱光愈敏感;相反,人在亮光处时,视紫红质的分解增强,合成过程甚弱,这就使视网膜中有较多的视紫红质处于分解状态,使视杆细胞几乎失去了感受光刺激的能力;事实上,人的视觉在亮光处是靠另一种对光刺激较不敏感的感光系统即视锥来完成的,后一系统在弱光时不足以被刺激,而在强光系统下视杆细胞中的视紫红质较多地处于分解状态时,视锥系统就代之而成为强光刺激的感受系统。在视紫红质和再合成的过程中,有一部分视黄醛被消耗,这最终要靠由食物进入血液循环(相当部分贮存于肝)中的维生素A来补充。长期摄入维生素A不足,将会影响人在暗光处的视力,引起夜盲症。
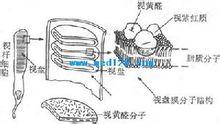
视杆细胞外段的超微结构示意图
视杆细胞外段的超微结构示意图感光细胞的外段是进行光-电转换的关键部位。视杆细胞外段具有特殊的超微结构。在外段部分,膜内的细胞浆甚少,绝大部分为一些整齐的重叠成层的圆盘状结构所占据,这圆盘称为视盘。每一个视盘是一个扁平的囊状物,囊膜的结构和细胞膜类似,具有一般的脂质双分子层结构,但其中镶嵌着的蛋白质绝大部分是视紫红质,亦即视杆细胞所含的视紫红质实际上几乎全部集中在视盘膜中。视盘的数目在不同动物的视杆细胞中相差很大,人的每个视杆细胞外段中它们的数目近千;每一个视盘所含的视紫红质分子约有100万个。这样的结构显然有利于使进入视网膜的光量子有更大的机会在外段中碰到视紫红质分子。

哺乳动物光感受器细胞模式
光子的吸收引起外段膜出现超极化电反应的机制已基本搞清,这就是光量子被作为受体的视紫红质吸收后引起视蛋白分子的变构,又激海参了视盘膜中一种称为传递蛋白(transducin)Ct的中介物,后者在结构上属于G-蛋白家庭的一员,它激活的结果是进而激活附近的磷酸二酯酶,于是使外段部分胞浆中的cGMP大量分解,而胞浆中cGMP的分解,就使未受光刺激时结合于外段膜的cGMP由也膜解离而被分解,而cGMP在膜上的存在正是这膜中存在的化学门控式Na+通道开放的条件,膜上cGMP减少,Na+通道开放减少,于是光照的结果出现了记录到的超极化型感受器电位。据估计,一个视紫红质被激活时,可使约500个传递蛋白被激活;虽然传递蛋白激活磷酸二酯酶是1对1的,但一个激活了的磷酸二酯酶在一秒钟内大约可使4千多个cGMP分子降解。由于酶系统的这种生物放大作用,就可以说明1个光量子的作用何以能在外段膜上引起大量化学门控式Na+通道的关闭,引起一个足以为人的视觉系统所感知的超极化型电变化。


