主要组织相容性复合体(具有免疫生理功能的基因群)

VLoG
次浏览
更新时间:2023-05-22
主要组织相容性复合体
具有免疫生理功能的基因群

主要组织相容性复合体( major histocompatibility complex,MHC) 是一组编码动物主要组织相容性抗原的基因群的统称。人类的MHC位于人的6号染色体短臂上,小鼠的MHC位于小鼠的17号染色体上。MHC的长度大约为4×10^6bp。人类的MHC也叫做HLA(human leukocyte antigen,HLA)复合体。小鼠的MHC则被称为H-2基因。由于MHC的多基因特性,依据其编码分子的结构、组织分布与功能差异,可分为MHC I类、MHC II类、MHC III类基因,分别编码MHC I类分子、MHC II类分子、MHC III类分子。人类的MHC产物通常被称为HLA (human leukocyte antigen,HLA),即人类白细胞抗原。
MHC的两个重要特性是其多基因性与高度多态性,由此造成每一种属的每一个体都有其特有的一套MHC分子。
发现MHC
在哺乳动物不同个体间进行皮肤移植时出现的排斥反应,具有记忆性、特异性和可转移性等免疫反应的基本特征,故从20世纪40年代起就确认移植排斥反应是一种典型的免疫现象。引起排斥反应的抗原称移植抗原(transplantation antigen)或组织相容性抗原(histocompatibility antigen)。此等抗原存在于细胞表面,无器官特异性,不同个体间其抗原特异性互不相同,但同卵双生及纯系动物不同个体之间,其抗原特异性完全一致。组织相容性抗原包括多种复杂的抗原系统。凡能引起快而强的排斥反应者称为主要组织相容性抗原系统,引起慢而弱的排斥反应者称为次要组织相容性抗原系统。若供者、受者双方的多个次要组织相容性抗原不匹配,同样会迅速发生明显的排斥反应。
现已证明,MHC不仅控制着同种移植排斥反应,更重要的是与机体免疫应答、免疫调节及某些病理状态的产生均密切相关。因此,MHC的完整概念是指脊椎动物某一染色体上编码主要组织相容性抗原、控制细胞间相互识别、调节免疫应答的一组紧密连锁基因群。
MHC分子
1、MHC分子的种类
不同的MHC编码的产物功能不同。
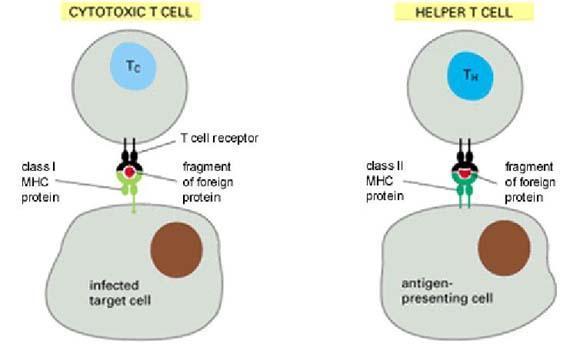
主要组织相容性复合体
MHC class Ⅱ(MHC Ⅱ):只位于抗原提呈细胞(APC)上,如巨噬细胞等。这类提供则是细胞外部的情况,像是组织中有细菌侵入,则巨噬细胞进行吞食后,把细菌碎片利用MHC提示给辅助T细胞,启动免疫反应。
MHC class Ⅲ (MHC Ⅲ) :主要编码补体成分,肿瘤坏死因子(TNF),热休克蛋白70(HSP70)和21羟化酶基因(CYP21A和CYP21B)。
2、MHC分子的生理意义
MHC抗原最初是作为移植抗原而被发现的,是引起移植排斥的主要抗原系统。这种抗原不合,即可引起受体的免疫应答,排斥移植的供体组织。70年代后证明MHC分子还具有重要的免疫生理功能。
MHC分子在免疫应答过程中参与抗原识别。70年代R.M.津克纳泽尔等在小鼠实验中发现杀伤 T细胞在杀伤感染病毒的靶细胞时,只能杀伤同系感染靶细胞,而对不同系的感染靶细胞则无杀伤作用,称这种现象为遗传限制性。随后证明杀伤T细胞与靶细胞的MHC必需一致才有杀伤作用,因此又称此现象为MHC限制性。
这揭示了MHC在T细胞识别异种抗原中的作用。进一步的研究证明T4的T细胞在识别异种抗原时受MHCⅡ类分子限制,而T8的T细胞识别异种抗原时受MHCⅠ类分子限制。这种限制性的机制是:T细胞通过其抗原识别受体,可同时识别异种抗原决定簇和自己MHC分子形成的新的复合抗原决定簇。
MHC基因
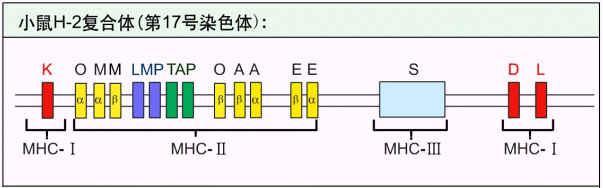
小鼠H-2复合体结构示意图
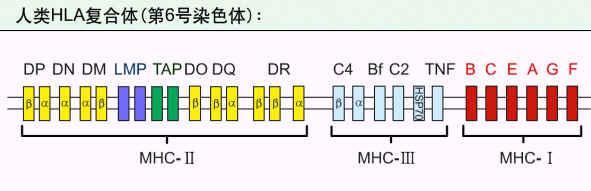
HLA复合体基因简图
1980年诺贝尔生理学或医学奖颁给了巴努·贝纳塞拉夫(Baruj Benacerraf)、吉罗格·D·斯奈尔(George D.Snell)和让·多塞(Jean Dausset,三人的研究为移植免疫学的确立奠定了基础。贝纳塞拉夫是美国医学家和免疫学家, 在研究器官移植排斥现象时, 发现了MHC(主要组织相容性复合体)中的免疫应答基因(Ir), 指出免疫现象由此基因所控制, 将免疫学在遗传学的基础上推向了高潮。
斯奈尔是美国免疫学家, 他通过对小鼠的组织移植实验提出: 不同个体间组织的可移植性是由细胞表面的特定抗原决定的, 即组织相容性抗原(也称H抗原), 由H基因控制。这种基因存在于某一染色体的有限区域, 这一区域被称为主要组织相容性复合体(MHC)。多塞是法国免疫学家, 他发现了人类白细胞抗原(HLA)和决定这些抗原的基因HLA基因, 即相当于小鼠的H基因; 还证实人类和其他许多动物都具有MHC。
MHC基因结构特点
例如:小鼠H-2:17号染色体:6号染色体短臂(6P21.31),全长3600-4000kb,224个基因座位(128个功
能基因,96个假基因)。
MHC的多态性的意义:
1 .扩大种群对抗原肽的提呈范围,有利于维持种群的生存与延续。
(HLA产物的多态性主要表现在抗原结合槽的氨基酸残基在组成和序列上不同)
2 .不利于器官移植中供体的选择。
HLA研究
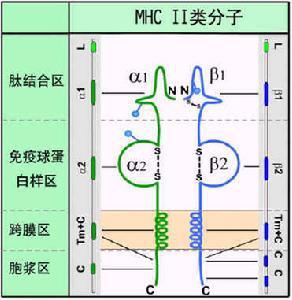
HLAⅠ类、Ⅱ类分子结构示意图
现在证明,在Ⅱ类基因区内存在与内源性抗原处理和递呈相关的基因,即LMP和TAP。LMP又称蛋白酶体相关基因(proteasome-related gene),由LMP2和LMP7两个基因组成,其编码产物LMP(low molecular mass polypeptide or large multifunctional protease)与内源性抗原的处理有关。TAP为多肽转运体基因,包括TAP1和TAP2两个基因,其编码产物TAP(transporter of antigenic peptides)与抗原肽的转运有关。HLA复合体Ⅰ类和Ⅱ类区基因名称见表5.1。
Ⅲ类基因区内已定位的至少有36个基因,其中与免疫系统有关的基因有C4B、C4A、C2、Bf、肿瘤坏死因子(TNFA、TNFB)和热休克蛋白70(HSP70),分别编码C4、C2、B因子、TNF-α、TNF-β和HSP70分子。在C4B两侧,还有与免疫系统无明显关系的CYP21B和CYP21A两个基因,编码21-羟化酶。大多数Ⅲ类基因产物合成后分泌到体液中去。具体内容见有关章节。HSP70主要在胞浆内,与其他蛋白质肽链的折叠、转位有关,亦可见于MΦ细胞和B细胞的内体(endosome)和膜表面,其作用为阻止内体中抗原的降解,并使之与Ⅱ类分子联合。
基因名称
HLA复合体Ⅰ类和Ⅱ类基因区内的基因名称
I类基因区 | Ⅱ类基因区 | ||
HLA-A | HLA-DRA | HLA-DQA1 | HLA-DRB |
HLA-B | HLA-DRB1 | HLA-DQB1 | HLA-DRB |
HLA-C | HLA-DRB2 | HLA-DQA2 | HLA-DRB |
HLA-E | HLA-DRB3 | HLA-DQB2 | HLA-DRB |
HLA-F | HLA-DRB4 | HLA-DQB3 |
展开表格
HLA与疾病
研究发现许多疾病与某些HLA等位基因或HLA单倍型确实呈现明显的相关性。与HLA抗原相关的疾病有几个应当注意的特点:病因和病理生理机理未明,以遗传模式分布但为弱的外显率;与免疫异常相关;对生殖影响很少或没有影响。
可用群体和家系研究来证实 HLA复合体内标记基因与各种疾病状态的相关性。因为群体研究易于进行,所以有关资料多来源于这种研究。具体疾病与某些具体HLA抗原的相关性通过计算相对风险性(RR)来定量,相对风险性可表述为具有疾病相关 HLA抗原的个体与缺乏这种抗原的个体相比发生该种疾病的机会。RR越高,在病人群体中该抗原的频率越高。以具有 HLA-B27的强直性脊椎炎病人为例。美国患该病的高加索人种90%具有HLA-B27,美国高加索人种对照者的具有率接近9%。RR=(90×91)÷(10×9)=91。因此,HLA-B27阳性个体发生该病的风险性是 HLA-B27阴性个体的91倍。因为不同种族之间某种抗原的频率通常有明显的不同。所以有必要在同种族中比较病人和对照者。例如,HLA-B27见于48%的患强直性脊椎炎的美国黑人,只见于 2%美国黑人对照者,则RR=45。
已经发现很多疾病与某种抗原相关。 HLA与疾病的相关性可分为统计学上较为肯定的相关性、可能相关性及潜在相关性几组。具肯定相关性的有:强直性脊椎炎(抗原为B27)、赖特尔氏综合征(B27)、急性前葡萄膜炎(B27)、青少年风湿性关节炎(B27)、乳糜泻(B8)、突眼性甲状腺肿(B8,DW3)、重症肌无力(B8,DR3)、疱疹性皮炎(B8)、慢性活动性肝炎(BW6,DR8)、青少年糖尿病(B8,DR3)、多发性硬化(B7,DR2)。具可能相关性的有:系统性红斑狼疮(DR3)、天疱疮(B13)、自身溶血性贫血(A3)、脊髓灰质炎(A3,B7)、贝赫切特氏病(B5)。具有潜在相关性的有:急性淋巴细胞白血病(A2,B12)、慢性肾小球肾炎(A2)、麻风(B14)、霍奇金氏病(A1,B8)。
参考资料
[1]
与免疫学有关的诺贝尔奖简介 · 360个人图书馆[引用日期2015-09-19]


