分子人类学(探究人类起源、民族演化的学科)

VLoG
次浏览
更新时间:2023-05-18

基本介绍
分子人类学(Molecular Anthropology)是人类学的分支,主要方法是比较DNA或蛋白质序列,早期方法亦包括血清学的比较研究。通过检查不同特定住民的DNA序列,科学家能判断特定住民之间或之内的亲属关系。人类学家们根据基因序列的特定相似处判断不同的人群是否属于同一基因组(haplogroup),以及是否发源自同一个地方。这就得以帮助人类学家跟踪迁徙和定居的模式,去发现现代人类如何形成和发展的。2002年,中国第一所研究分子人类学的学术机构复旦大学现代人类学研究中心正式成立。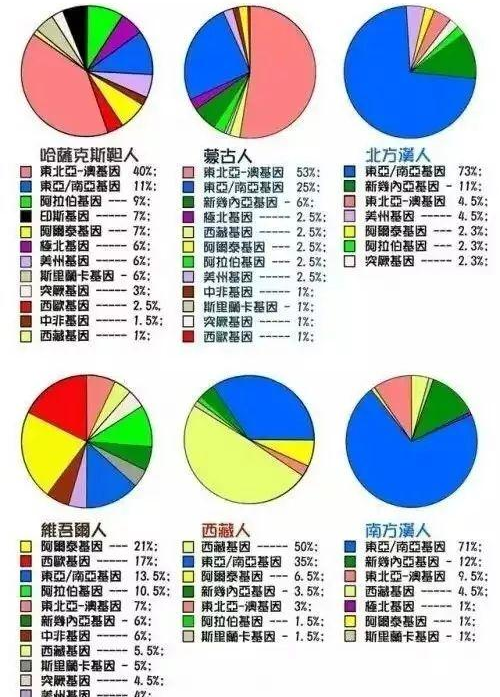

分子人类学
在过去分子人类学还没有兴起的年代,研究历史探寻民族起源只能靠有限的史料和考古发现来大概推测,这是非常具有局限性的。尤其是史料,是非常有限的,也未必是真实的。所以,分子人类学的兴起将大大打破这些局限,向人们揭示一个更真实更准确的历史,从而更加清晰地分析一个民族的起源。分子人类学用DNA材料和计算生物学方法解答了很多人类学的问题。
与人类起源论
分子人类学技术大量应用于人类学和考古学研究领域,在人类起源与演化、人群的迁徙与交流、人群间的亲缘关系,以及考古鉴定等方面发挥着越来越大的作用。1987年,英国《自然》(Nature)周刊上刊登了美国加州大学伯克利分校三位分子生物学家卡恩(R. L. Cann)、斯通金(M. Stoneking)和威尔逊(A. C. Wilson)的《线粒体DNA与人类进化》(Mitochondrial DNA and human evolution)一文,他们选择了其祖先来自非洲、欧洲、亚洲、中东,以及巴布亚新几内亚和澳大利亚土著共147名妇女,从她们生产后婴儿的胎盘细胞中成功地提取出mtDNA,并对其序列进行了分析,根据分析结果绘制出一个系统树。由此推测,所测定的婴儿mtDNA可以将所有现代人最后追溯到大约29万~14万年,平均20万年前生活在非洲的一位妇女。她就是今天生活在地球上各个角落的人的共同“祖母”。其后,又根据mtDNA发生突变的速率计算出非洲人群分化出世界其他人群的大致时间,为大约18万~9万年,平均约13万年前。认为在大约13万年前,这个“祖母”的一群后裔离开了他们的家园非洲,向世界各地迁徙扩散,并逐渐取代了生活在当地的土著居民直立人的后裔早期智人,从此在世界各地定居下来,逐渐演化发展成现在的我们。这就是著名的现代人起源的“夏娃假说”。
2000年美国斯坦福大学昂德希尔(P. A. Underhill)等利用变性高效液相层析技术,分析得到218个Y染色体非重组区位点构成的131个单倍型,对全球1062个具有代表性的男性个体进行研究,同样根据分析结果绘制出一个系统树。Y-DNA系统树所展示的结果与mtDNA系统树的结果非常相似。欧洲和亚洲等世界其他现代人群都起源于非洲,而美洲和澳洲现代人群又都起源于亚洲人群。这就是与“夏娃假说”相互应征的“亚当假说”。同样根据Y-DNA发生突变的速率计算出非洲人群分化出世界其他人群的大致时间在14万~4万年,平均约6万年前。
1997年7月,美国《科学》(Science)周刊发表了一篇文章,引起学术界一片喧嚣。德国慕尼黑大学的分子生物学家克林斯(M. Krings)等,对1856年发现于德国杜塞尔多夫城尼安德特峡谷的距今大约6万年左右的尼安德特人化石,进行了mtDNA的抽提和PCR扩增,并对提取出的DNA进行了测序。发现尼人的mtDNA序列中有12个片断与现代人类的完全不同,尼人的mtDNA处在现代人类的变异范围之外,推算得出的分化时间在30万年以上。而历史上尼人和现代人的并存历史在10万年以内,如果这两个人种之间有直接传承关系,其差异应该不超过10万年。由此推测,尼人不可能是现代人类的直系祖先,他们根本就没有将其血缘遗传给现代人类,只成为人类演化史上的一个旁支。这一研究结果支持“现代人起源于非洲的假说”。这一科学发现曾被评为1997年世界十大科技成就之一。其后,又有科学家成功地从出土于高加索和克罗地亚的尼安德特人化石中提取了mtDNA序列,同样得出尼人与现代人没有遗传联系的结论。
褚嘉佑等14位中国学者1998年在《美国科学院学报》(Proceedings of the National Academy of Sciences of the United Sates of America,PNAS)上发表了一篇文章也支持现代人起源于非洲的观点。他们利用30个常染色体微卫星位点(由2-6个碱基重复单位构成的DNA序列),分析了包括中国汉族和少数民族的南北人群在内的28个东亚人群的遗传结构,结果支持现代中国人也起源于非洲的假说。并且认为现代中国人群是由东南亚进入中国大陆,而非通过中亚移民过来的。但是由于样本量较少、群体代表性不强,且微卫星位点突变率较高,对追溯久远事件有一定局限性等原因,褚嘉佑等人的工作对证明东亚人群起源于非洲的观点还不十分令人信服。
1999年,宿兵等人对包括中国各省份的汉族和少数民族,以及东北亚、东南亚、非洲、美洲和大洋洲总共925个个体的不同人群,利用19个Y-SNP(Y染色体单核苷酸多态位点)构成的一组Y染色体单倍型,系统地研究了包括中国各人群在内的现代东亚人的起源和迁徙。结果显示包括中国各人群在内的所有现代东亚人群的Y-SNP单倍型均来自较晚发生的突变,而更早的类型仅存在于非洲。因此认为,现代东亚人全部来自于非洲的某个古代类型。而且,东亚人群的迁徙是从东南亚进入到中国的南方,再向北迁移逐渐扩散到中国各地区及东北亚,并有可能完全取代了生活在当地的原土著居民,而成为该地区的新居民。
对于经过漫长跋涉由非洲迁徙而来的这些现代人群,是完全取代了生活在东亚大陆上的原土著居民成为东亚地区的新居民,还是与当地的土著居民有着某些程度的融合,两者共同对现在的我们有遗传贡献呢?这个问题也令许多科学家有非常浓厚的兴趣。他们基于这样一个假设,即从非洲迁徙而来的现代人群并没有在群体上完全取代当地土著居民,当地土著居民也可能有少量的基因遗传下来,在以上实验的基础上扩大东亚地区的样本量,来看这些当地土著居民的可能贡献率或不完全取代的可能性有多少?这些遗留下来的基因又有可能保留在现代哪些人群中?
2001年,柯越海等人对来自中国各地区近12 000份男性随机样本进行了M89、M130和YAP三个Y染色体单倍型的分型研究。所选择的三个Y染色体非重组区的突变型M89、M130和YAP均来自另一个Y染色体单倍型M168。M168突变型是人类走出非洲并扩散到非洲以外其他地区的代表性突变位点,它是所有非洲以外人群Y染色体的最近的共同祖先,所以M168是现代人类单一起源于非洲的最直接证据,在除非洲以外的其他地区没有发现一例个体具有比M168更古老的突变型。该项研究结果显示万份样品无一例外具有M89、M130和YAP三种突变型之一,并没有发现个体携带有以上三种Y-SNP突变型之外的类型,也没有发现同时具有M89、M130和YAP突变中任意两个以上突变的个体,这一结果与非洲以外的世界其他地区的基因型分型结果是一致的。在所检测的所有中国12000份样品中全部都携带有来自非洲的M168突变型的“遗传痕迹”,因此认为,Y染色体的证据并不支持中国现代人独立起源的假说,而支持包括中国人在内的东亚现代人起源于非洲的假说。
单倍体遗传点
男性和女性携带有两套连续遗传机制。第一套是Y染色体在男性间的遗传,从父到子。第二套连续遗传机制是线粒体DNA,即mtDNA在女性之间进行遗传。只有非常特殊的情况下,mtDNA在男性间进行遗传。
线粒体DNA
线粒体存在于细胞质里,是细胞的“能量工厂”,它们包含少量遗传物质---DNA。每个人的线粒体都来自母亲。与作为基因组的染色体DNA不同,线粒体DNA的优点在于它不会进行重组。DNA重组过于频繁就丧失追溯到父系源流的能力。而线粒体DNA却是在不断克隆自身,只会接受极少的父系mtDNA。线粒体DNA不但所处的位置与细胞核DNA不同,遗传方式也迥然有别--它测定线粒体DNA是追踪母系血亲的惟一办法。线粒体DNA的另一点优势在于,高度变化区域的进化快速,显示线粒体DNA的某些染色体领域趋于局外中立。因此线粒体DNA还被当作一种特殊的“钟”,用来估计家系大约是在多久之前彼此分离的。因此线粒体是从母系遗传的角度研究人类进化的重要工具,就像Y染色体是研究父系遗传的工具。同时,线粒体基因组是独立于核基因组的遗传物质,它普遍存在于真核细胞中,线粒体内包含有DNA和转录与转译系统,是具有一定自主性的细胞器。线粒体基因组具有的独特优点:线粒体DNA分子小、拷贝数高; 结构和组织简单而高度保守; 母系遗传,缺乏重组; DNA突变率高。线粒体DNA跟RFLP的原理是一样的,只不过目的DNA由基因组DNA变为线粒体DNA。
除男性精子细胞外,人身体所有细胞里面都有线粒体,但只有女性的线粒体基因能随其卵子遗传给后代。mtDNA是Mitochondrial DNA(线粒体DNA)的缩写,是承载线粒体遗传密码的物质。男人线粒体只伴随此男人生活一生,然后终结,不能遗传给后代。mtDNA表现为母系遗传。mtDNA结构类型是反映母系脉络的重要指标。通过检测现代人mtDNA,能弄清各民族、各地人的母系血缘关系。通过检测古尸线粒体,可弄清历史上各个民族间的母系血缘关系、历史故事、迁徙路线、历史名人的民族、身份。
母亲给儿女贡献了50%的遗传基因,对儿女遗传特性有着和父亲一样的影响力。古代一夫多妻,有时抢掠战败民族女性为妻为妾。胜族由于不适应败族地区气候地理条件,一般打完胜仗带着抢来的妾返回祖籍并共同生活生育儿女。异族妻妾生育的孩子虽有一半异族基因,但文化上被视作其父民族的人。儿子长大后继续到异族领地抢掠妻妾,生育的孙子辈已有75%异族血统,但仍被视作属于其爷爷民族的人。虽然孙子的Y染色体仍然和爷爷的一样类型,但其母系线粒体mtDNA,以及身体常染色体已和其爷爷的大不相同。如此不断循环,导致民族的文化、语言虽然还是祖先的,但若干代后民族人口的血统、基因已发生很大改变。
因此,一个民族、地区人群的母系线粒体mtDNA结构类型和构成比例,比父系Y染色体更能反映其遗传和血统特性。
Y染色体
Y染色体存在于细胞核中,即细胞核DNA(nDNA)。与mtDNA不同的是,Y染色体在非重组区域存在交换。
基因的本质是DNA链上有功能的片断,它参与蛋白质等的合成,最终决定了生物的特征,它的多样性也决定了生物界的多样性。此外,DNA链上还有更多没有具体表达功能的片断,这些片断不受自然选择压力的影响,可以匀速自由地突变,更忠实地记载了人群进化的历史。
DNA遗传
线粒体只会遗传自母亲,以哺乳类而言,一般在受精之后,卵子细胞就会将精子中的线粒体摧毁。 1999年发表的研究中显示,父系精子线粒体(含有mtDNA)带有泛素(ubiquitin)标记,因而在胚胎中会被挑选出来,进而遭到摧毁。不过某些细胞外的人工受精技术可直接将精子注入卵子细胞内,可能会干扰摧毁精子线粒体的过程。
由于mtDNA并非高度保守,而是拥有较快的突变速率,因此可用来研究种系发生学,生物学家挑选少量不同物种的基因,分析其序列的保留与变异程度,可建立出演化树。
假设
不可靠性重组
科学家认为,线粒体DNA分子是相对稳定的,不会互相交换DNA片断,造成它们发生变化的唯一因素是自发变异。这种变异以相对稳定的速率进行并积累,可以作为“分子钟”使用。两个人的线粒体DNA的差异程度,就决定了这两个人最近的母系共同祖先生活在多少年前。科学家曾经对世界不同地区和民族的女性进行线粒体DNA调查,确定现代人的线粒体来自于约15万年前的一位女性,这位母系祖先被称为“线粒体夏娃”。但在几年前,人们发现了一个罕见的例外,一名男子的线粒体DNA中,有一部分来自于他的父亲。为了检验线粒体DNA不会重组的假设是否正确,哈佛医学院的科学家对这名男子和他的父母进行线粒体DNA序列比较。结果发现,有一些来自父亲的线粒体DNA片断混合在母系DNA中。进一步试验表明,负责复制线粒体DNA的酶停止复制母亲的DNA、跳到父亲的DNA上从对应的位置开始复制时,就发生了线粒体DNA的重组。这一成果显示,线粒体DNA可能并不那么稳定,而会因为自发变异之外的原因发生改变。寻找人类母系祖先的研究,比人们原先认为的更复杂。
汉族先民
根据2007年研究,山西陶寺遗址的龙山文化居民中,上层贵族100%为O3类型,而根据2008年金力、李辉的数据,陕西仰韶文化的居民的Y染色体SNP类型也是O3,所以,我们可以清晰的看出,古代汉族先民,尤其是贵族,都是O3占有绝对主体的,仰韶文化和龙山文化居民的Y染色体成分是基本相同的,种族上是同一成分。
O3是汉族的绝对主体,占据非常重要的位置,根据2004年文波的数据,山东汉族为59.5%(110/185)、河南52%(26/50)、陕西汉族56.7%(51/90)、云南汉族63.4%(59/93)、广东汉族53.9%(34/64)、四川汉族57.1%(36/63)、浙江汉族是50.9%(54/106),张咏莉2002年福建汉族的为58.8%(47/80),2006年Hammer的数据北方汉族65.9%(29/44)、台湾汉族49/84(58.3%)(注意这个数据和张咏莉2002年福建汉族的58.8数据几乎一样,类似的还有Nonaka的台湾汉族,183个里106个O3,比例58.2%,福建闽南汉族和台湾汉族数据高度一致),XUE的2006年数据,哈尔滨汉族O3频率是68.6%(23/35),Lu chuncheng的数据中南京汉族为53.6%(429/800)、马明义的数据中四川汉族61.8%(211/341)2007年Zhou ruixia的数据,甘肃71.3%(62/87),所有数据中,除了河北人83.3%(10/12)由于样本太少外,根据2003年李辉的数据显示,福建长汀客家人中O3的比例高达74.3%(110/148)。还有一个类似的数据,是2002年李辉在《广西六甲人来源》一文中的数据,为77.8%,(21人/27人)。
总体上看,所有汉族中,O3都是主体的,不同的取样可能造成数据有波动,但总体出现频率大部分一般在50-70%之间,虽然幅员辽阔人口众多,但汉族内部却表现出了惊人的一致,显示了汉族的共同起源。
今天12亿汉族的绝大多数,都是5000年前中原原始居民的直系后代。主体是仰韶文化和龙山文化的O3(两种文化居民的父系成分相同),O3也就是来自中原仰韶文化和龙山文化的居民构成了各地汉族父系远祖的压倒性绝对多数,其次是河北的磁山文化的O1,红山文化的O3、O2和O1,和良渚文化的O1,其次还有湖北大溪文化和江西吴城文化的O2a,今天广阔的中国土地,仍然为5000年前中国远古汉族先民的子孙所占据。
随着研究不断深入,南线学说无法全面解释现有东亚人群南北方的遗传差异,以及北部地区检测到的属于中亚和欧洲地区的遗传信号。对于“北线”假说是否成立,以及中亚及欧洲遗传信号是否仅仅由于近期(而非史前)基因交流的结果,在之前缺少有力证据支持。
研究表明,东亚人群中存在4个主要Y-单倍群占检测人数约93%,支持南线学说。重要的是,研究发现东亚确实存在中亚和西部欧亚的遗传输入(约7%)。对这些单倍群分别进行系统地理发育分析发现,它们基本都表现出沿北线的末次冰川期后的人群扩张。尤其是Y染色体单倍群Q的一个亚分支Q1a1-M120,推测是史前人群沿北线迁移,途径东亚北部留下的最好痕迹。Y染色体单倍群Q分布在东欧、北欧、高加索、西亚、南亚、中亚、西伯利亚、美洲和东亚,但Q的分支Q1a1-M120局限性地分布在东亚,主要是东亚北方人群,并且表现出了最高的多态性。另外,美洲的Q来源人群(Q1a3a)的祖先单倍群Q1a3*在东亚北部也有检出。单倍群Q1a1-M120和Q1a3*-M346来源的个体表现出的共祖时间分别是1.54和1.78万年。这些证据很好地支持了史前人群走北线的迁移方式,并且正是这种北线对东亚的人口输入,加大了东亚南北方人群的遗传差异。
人口移动
Genetic Evidence Supports Demic Diffusion of Han Culture,(“遗传学证实汉文化的扩散源于人口扩张”)(Bo WEN, Hui LI, et al, Nature, No.431,September 2004, pp.302 - 305)
(英国的《自然》杂志,与美国的《科学》和《细胞》杂志被公认为世界自然科学界三个最顶级的学术刊物。)论文用基因学证据,论证了南北方汉族父系血统的相同起源,汉文化的扩散源于人口扩张,而不是所谓的"融合"。南北汉族的相貌体格的差异主要是母系差异和地理环境造成的。
南方汉族
据史料记载和考古发现,汉族起源于黄河流域,是从黄河流域不断往南迁徙的。根据复旦大学的基因研究对照历史迁移记录,汉民族的扩张主因是历史上的由北往南的人口移动,并非所谓的南方少数民族接受汉文化形成了南方汉族。南方汉族的父系近八成来自中原南迁的北方汉族,母系则近六成来自南迁的北方汉族。也就是说现代南方汉族主要成分是从中原南迁的北方汉族,而不是所谓的南方少数民族接受汉文化而形成了南方汉族。


