脱氧核糖核酸(主要的遗传物质)

VLoG
次浏览
更新时间:2023-05-17
脱氧核糖核酸
主要的遗传物质
人体内重要的遗传物质
基本信息
| 中文名 | 脱氧核糖核酸 |
| 外文名 | deoxyribonucleic acid |
| 功能 | 长期性的信息储存 |
| 复制方式 | 半保留复制 |
| 分子结构 | 双螺旋结构 |
展开
简介
脱氧核糖核酸(DNA)是生物细胞内携带有合成RNA和蛋白质所必需的遗传信息的一种核酸。
历史沿革
组成与功能
20世纪初,德国科赛尔(1853-1927)和他的两个学生琼斯(1865-1935)和列文(1869-1940)的研究,弄清了核酸的基本化学结构,认为它是由许多核苷酸组成的大分子。核苷酸是由碱基、核糖和磷酸构成的。其中碱基有4种(腺嘌呤、鸟嘌呤、胸腺嘧啶和胞嘧啶),核糖有两种(核糖、脱氧核糖),因此把核酸分为核糖核酸(RNA)和脱氧核糖核酸(DNA)。
列文急于发表他的研究成果,错误地认为4种碱基在核酸中的量是相等的,从而推导出核酸的基本结构是由4个含不同碱基的核苷酸连接成的四核苷酸,以此为基础聚合成核酸,提出了“四核苷酸假说”。这个错误的假说,对认识复杂的核酸结构起了相当大的阻碍作用,也在一定程度上影响了人们对核酸功能的认识。
1902年,德国化学家费歇尔提出氨基酸之间以肽链相连接而形成蛋白质的理论,1917年他合成了由15个甘氨酸和3个亮氨酸组成的18个肽的长链。于是,有的科学家设想,很可能是蛋白质在遗传中起主要作用。如果核酸参与遗传作用,也必然是与蛋白质连在一起的核蛋白在起作用。因此,那时生物界普遍倾向于认为蛋白质是遗传信息的载体。
到了1919年,菲巴斯·利文进一步辨识出组成DNA的碱基、糖类以及磷酸核苷酸单元,他认为DNA可能是许多核苷酸经由磷酸基团的联结,而串联在一起。不过他所提出概念中,DNA长链较短,且其中的碱基是以固定顺序重复排列。1937年,威廉·阿斯特伯里完成了第一张X光绕射图,阐明了DNA结构的规律性。
脱氧核糖核酸
双螺旋的发现
20世纪30年代后期,瑞典的科学家们就证明DNA是不对称的。第二次世界大战后,用电子显微镜测定出DNA分子的直径约为2nm。DNA双螺旋结构被发现后,极大地震动了学术界,启发了人们的思想。从此,人们立即以遗传学为中心开展了大量的分子生物学的研究。首先是围绕着4 种碱基怎样排列组合进行编码才能表达出20种氨基酸为中心开展实验研究。
20世纪50年代,DNA双螺旋结构被阐明,揭开了生命科学的新篇章,开创了科学技术的新时代。随后,遗传的分子机理――DNA复制、遗传密码、遗传信息传递的中心法则、作为遗传的基本单位和细胞工程蓝图的基因以及基因表达的调控相继被认识。至此,人们已完全认识到掌握所有生物命运的东西就是DNA和它所包含的基因,生物的进化过程和生命过程的不同,就是因为DNA和基因运作轨迹不同所致。
1953年4月25日,英国的《自然》杂志刊登了美国的沃森和英国的克里克在英国剑桥大学合作的研究成果:DNA双螺旋结构的分子模型,这一成果后来被誉为20世纪以来生物学方面最伟大的发现,标志着分子生物学的诞生。

在大学期间,沃森在遗传学方面虽然很少有正规的训练,但自从阅读了薛定谔的《生命是什么?--活细胞的物理面貌》这本进化论的理论基础书籍,促使他去“发现基因的秘密”。他善于集思广益,博取众长,善于用他人的思想来充实自己。只要有便利的条件,不必强迫自己学习整个新领域,也能得到所需要的知识。

克里克
当时克里克比沃森大12岁,还没有取得博士学位。但他们谈得很投机,沃森感到在这里居然能找到一位懂得DNA比蛋白质更重要的人,真是三生有幸。同时沃森感到在他所接触的人当中,克里克是最聪明的一个。他们每天交谈至少几个小时,讨论学术问题。两个人互相补充,互相批评以及相互激发出对方的灵感。
他们认为解决DNA分子结构是打开遗传之谜的关键。只有借助于精确的X射线衍射资料,才能更快地弄清DNA的结构。为了搞到DNAX射线衍射资料,克里克请威尔金斯到剑桥来度周末。在交谈中威尔金斯接受了DNA结构是螺旋型的观点,还谈到他的合作者富兰克林(1920--1958,女)以及实验室的科学家们,也在苦苦思索着DNA结构模型的问题。从1951年11月至1953年4月的18个月中,沃森、克里克同威尔金斯、富兰克林之间有过几次重要的学术交往。
1951年11月,沃森听了富兰克林关于DNA结构的较详细的报告后,深受启发,具有一定晶体结构分析知识的沃森和克里克认识到,要想很快建立 DNA结构模型,只能利用别人的分析数据。他们很快就提出了一个三股螺旋的DNA结构的设想。1951年底,他们请威尔金斯和富兰克林来讨论这个模型时,富兰克林指出他们把DNA的含水量少算了一半,于是第一次设立的模型宣告失败。
有一天,沃森又到国王学院威尔金斯实验室,立刻兴奋起来、心跳也加快了,因为这种图像比以前得到的“A型”简单得多,只要稍稍看一下“B型”的X射线衍射照片,再经简单计算,就能确定DNA分子内多核苷酸链的数目了。
他们苦苦地思索4种碱基的排列顺序,一次又一次地在纸上画碱基结构式,摆弄模型,一次次地提出假设,又一次次地推翻自己的假设。
有一次,沃森又在按着自己的设想摆弄模型,他把碱基移来移去寻找各种配对的可能性。突然,他发现由两个氢键连接的腺嘌呤一胸腺嘧啶对竟然和由3个氢键连接的鸟嘌呤一胞嘧啶对有着相同的形状,于是精神为之大振。因为嘌呤的数目为什么和嘧啶数目完全相同这个谜就要被解开了。查伽夫规律也就一下子成了 DNA双螺旋结构的必然结果。因此,一条链如何作为模板合成另一条互补碱基顺序的链也就不难想象了。那么,两条链的骨架一定是方向相反的。
经过沃森和克里克紧张连续的工作,很快就完成了DNA金属模型的组装。从这模型中看到,DNA由两条核苷酸链组成,它们沿着中心轴以相反方向相互缠绕在一起,很像一座螺旋形的楼梯,两侧扶手是两条多核苷酸链的糖一磷基因交替结合的骨架,而踏板就是碱基对。由于缺乏准确的X射线资料,他们还不敢断定模型是完全正确的。
下一步的科学方法就是把根据这个模型预测出的衍射图与X射线的实验数据作一番认真的比较。他们又一次打电话请来了威尔金斯。不到两天工夫,威尔金斯和富兰克林就用X射线数据分析证实了双螺旋结构模型是正确的,并写了两篇实验报告同时发表在英国《自然》杂志上。1962年,沃森、克里克和威尔金斯获得了诺贝尔医学和生理学奖,而富兰克林因患癌症于1958年病逝而未被授予该奖。
理化性质
脱氧核糖核酸
腺嘌呤脱氧核苷酸(dAMP )、胸腺嘧啶脱氧核苷酸(dTMP )、胞嘧啶脱氧核苷酸(dCMP )、鸟嘌呤脱氧核苷酸(dGMP )。
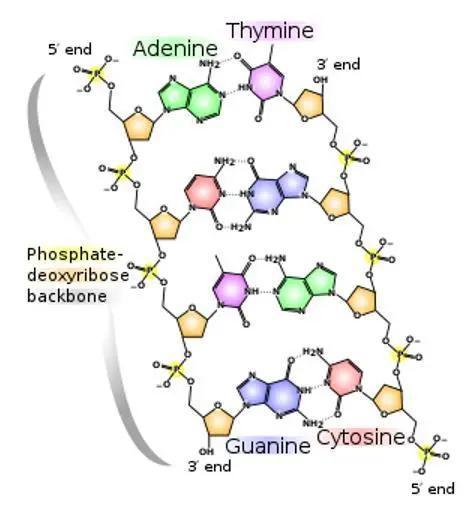
脱氧核糖核酸是一种由核苷酸重复排列组成的长链聚合物,宽度约22到24埃(2.2到2.4纳米),每一个核苷酸单位则大约长3.3埃(0.33纳米)。在整个脱氧核糖核酸聚合物中,可能含有数百万个相连的核苷酸。例如人类细胞中最大的1号染色体中,就有2亿2千万个碱基对。通常在生物体内,脱氧核糖核酸并非单一分子,而是形成两条互相配对并紧密结合,且如藤蔓般地缠绕成双螺旋结构的分子。每个核苷酸分子的其中一部分会相互连结,组成长链骨架;另一部分称为碱基,可使成对的两条脱氧核糖核酸相互结合。所谓核苷酸,是指一个核苷加上一个或多个磷酸基团,核苷则是指一个碱基加上一个糖类分子。
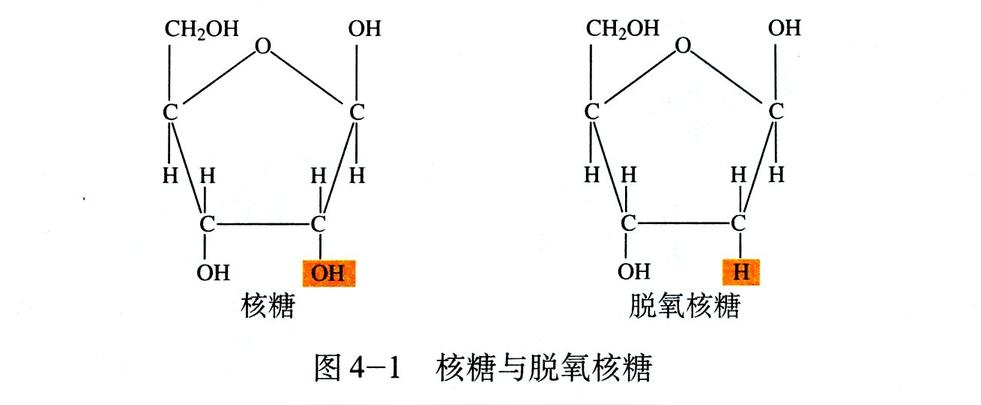
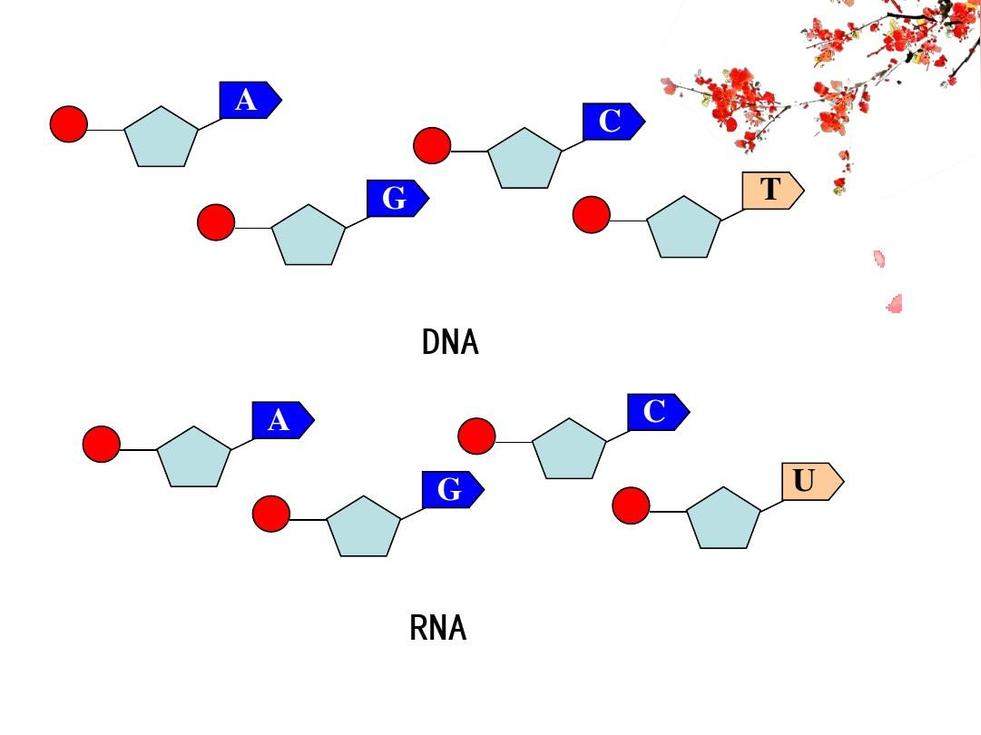
脱氧核糖核酸骨架是由磷酸与糖类基团交互排列而成。组成脱氧核糖核酸的糖类分子为环状的2-脱氧核糖,属于五碳糖的一种。磷酸基团上的两个氧原子分别接在五碳糖的3号及5号碳原子上,形成磷酸双酯键。这种两侧不对称的共价键位置,使每一条脱氧核糖核酸长链皆具方向性。双螺旋中的两股核苷酸互以相反方向排列,这种排列方式称为反平行。脱氧核糖核酸链上互不对称的两末端一边叫做5'端,另一边则称3'端。脱氧核糖核酸与RNA最主要的差异之一,在于组成糖分子的不同,DNA为2-脱氧核糖,RNA则为核糖。
而脱氧核糖(五碳糖)与磷酸分子借由酯键相连,组成其长链骨架,排列在外侧,四种碱基排列在内侧。每个糖分子都与四种碱基里的其中一种相连,这些碱基沿着DNA长链所排列而成的序列,可组成遗传密码,指导蛋白质的合成。读取密码的过程称为转录,是以DNA双链中的一条单链为模板转录出一段称为mRNA(信使RNA)的核酸分子。多数RNA带有合成蛋白质的讯息,另有一些本身就拥有特殊功能,例如rRNA、snRNA与siRNA。
在细胞内,DNA能与蛋白质结合形成染色体,整组染色体则统称为染色体组。对于人类而言,正常的人体细胞中含有46条染色体。染色体在细胞分裂之前会先在分裂间期完成复制,细胞分裂间期又可划分为:G1期-DNA合成前期、S期-DNA合成期、G2-DNA合成后期。对于真核生物,如动物、植物及真菌而言,染色体主要存在于细胞核内;而对于原核生物,如细菌而言,则主要存在于细胞质中的拟核内。染色体上的染色质蛋白,如组织蛋白,能够将DNA进行组织并压缩,以帮助DNA与其他蛋白质进行交互作用,进而调节基因的转录。
DNA是高分子聚合物,DNA溶液为高分子溶液,具有很高的粘度,可被甲基绿染成绿色。DNA对紫外线(260nm)有吸收作用,利用这一特性,可以对DNA进行含量测定。当核酸变性时,吸光度升高,称为增色效应;当变性核酸重新复性时,吸光度又会恢复到原来的水平。较高温度、有机溶剂、酸碱试剂、尿素、酰胺等都可以引起DNA分子变性,即DNA双链碱基间的氢键断裂,双螺旋结构解开—也称为DNA的解螺旋。
分子结构
DNA是由许多脱氧核苷酸按一定碱基顺序彼此用3’,5’-磷酸二酯键相连构成的长链。大多数DNA含有两条这样的长链,也有的DNA为单链,如大肠杆菌噬菌体φX174、G4、M13等。DNA有环形DNA和链状DNA之分。在某些类型的DNA中,5-甲基胞嘧啶可在一定限度内取代胞嘧啶,其中小麦胚DNA的5-甲基胞嘧啶特别丰富。在某些噬菌体中,5-羟甲基胞嘧啶取代了胞嘧啶。40年代后期,查伽夫(E.Chargaff)发现不同物种DNA的碱基组成不同,但其中的腺嘌呤数等于其胸腺嘧啶数(A=T),鸟嘌呤数等于胞嘧啶数(G=C),因而嘌呤数之和等于嘧啶数之和,一般用几个层次描绘DNA的结构。
一级结构
是指构成核酸的四种基本组成单位——脱氧核糖核苷酸(核苷酸),通过3',5'-磷酸二酯键彼此连接起来的线形多聚体,以及其基本单位-脱氧核糖核苷酸的排列顺序。 每一种脱氧核糖核苷酸由三个部分所组成:一分子含氮碱基+一分子五碳糖(脱氧核糖)+一分子磷酸根。核酸的含氮碱基又可分为四类:腺嘌呤(adenine,缩写为A),胸腺嘧啶(thymine,缩写为T),胞嘧啶(cytosine,缩写为C)和鸟嘌呤(guanine,缩写为G)。DNA的四种含氮碱基组成具有物种特异性。即四种含氮碱基的比例在同物种不同个体间是一致的,但在不同物种间则有差异。DNA的四种含氮碱基比例具有奇特的规律性,每一种生物体DNA中 A=T ,C=G 查伽夫规则(即碱基互补配对原则)。
每一种脱氧核糖核苷酸由三个部分所组成:一分子含氮碱基+一分子五碳糖(脱氧核糖)+一分子磷酸根。核酸的含氮碱基又可分为四类:腺嘌呤(adenine,缩写为A),胸腺嘧啶(thymine,缩写为T),胞嘧啶(cytosine,缩写为C)和鸟嘌呤(guanine,缩写为G)。DNA的四种含氮碱基组成具有物种特异性。即四种含氮碱基的比例在同物种不同个体间是一致的,但在不同物种间则有差异。DNA的四种含氮碱基比例具有奇特的规律性,每一种生物体DNA中 A=T ,C=G 查伽夫规则(即碱基互补配对原则)。

脱氧核糖核酸
脱氧核糖核酸
三级结构
是指DNA中单链与双链、双链之间的相互作用形成的三链或四链结构。如H-DNA或R-环等三级结构。DNA的三级结构是指DNA进一步扭曲盘绕所形成的特定空间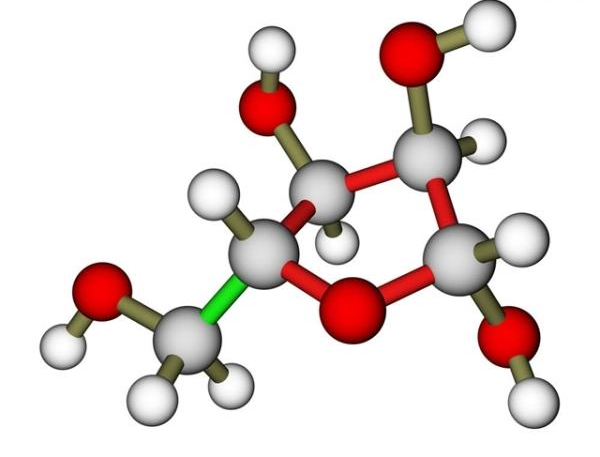 结构,也称为超螺旋结构。DNA的超螺旋结构可分为正、负超螺旋两大类,并可互相转变。超螺旋是克服张力而形成的。当DNA双螺旋分子在溶液中以一定构象自由存在时,双螺旋处于能量最低状态此为松弛态。如果使这种正常的DNA分子额外地多转几圈或少转几圈,就是双螺旋产生张力,如果DNA分子两端是开放的,这种张力可通过链的转动而释放出来,DNA就恢复到正常的双螺旋状态。但如果DNA分子两端是固定的,或者是环状分子,这种张力就不能通过链的旋转释放掉,只能使DNA分子本身发生扭曲,以此抵消张力,这就形成超螺旋,是双螺旋的螺旋。
结构,也称为超螺旋结构。DNA的超螺旋结构可分为正、负超螺旋两大类,并可互相转变。超螺旋是克服张力而形成的。当DNA双螺旋分子在溶液中以一定构象自由存在时,双螺旋处于能量最低状态此为松弛态。如果使这种正常的DNA分子额外地多转几圈或少转几圈,就是双螺旋产生张力,如果DNA分子两端是开放的,这种张力可通过链的转动而释放出来,DNA就恢复到正常的双螺旋状态。但如果DNA分子两端是固定的,或者是环状分子,这种张力就不能通过链的旋转释放掉,只能使DNA分子本身发生扭曲,以此抵消张力,这就形成超螺旋,是双螺旋的螺旋。

脱氧核糖核酸
四级结构
拓扑结构
也是DNA存在的一种形式。DNA的拓扑结构是指在DNA双螺旋的基础上,进一步扭曲所形成的特定空间结构。超螺旋结构是拓扑结构的主要形式,它可以分为正超螺旋和负超螺旋两类,在相应条件下,它们可以相互转变。
其他结构
主要类别
单链DNA
单链DNA(single-stranded DNA)大部分DNA以双螺旋结构存在,但一经热或碱处理就会变为单链状态。单链DNA就是指以这种状态存在的DNA。单链DNA在分子流体力学性质、吸收光谱、碱基反应性质等方面都和双链DNA不同。某些噬菌体粒子内含有单链环状的DNA,这样的噬菌体DNA在细胞内增殖时则形成双链DNA。
闭环DNA
闭环DNA(closed circular DNA)没有断口的双链环状DNA,亦称为超螺旋DNA。由于具有螺旋结构的双链各自闭合,结果使整个DNA分子进一步旋曲而形成三级结构。另外如果一条或二条链的不同部位上产生一个断口,就会成为无旋曲的开环DNA分子。从细胞中提取出来的质粒或病毒DNA都含有闭环和开环这二种分子。可根据两者与色素结合能力的不同,而将两者分离开来。
连接DNA
连接DNA (Linker DNA):核小体中除147bp核心DNA 外的所有DNA。
模板DNA
模板DNA可以是单链分子,也可以是双链分子,可以是线状分子,也可以是环状分子(线状分子比环状分子的扩增效果稍好)。就模板DNA而言,影响PCR的主要因素是模板的数量和纯度。
互补DNA
互补DNA(cDNA,complementary DNA)构成基因的双链DNA分子用一条单链作为模板,转录产生与其序列互补的信使RNA分子,然后在反转录酶的作用下,以mRNA分子为模板,合成一条与mRNA序列互补的单链DNA,最后再以单链DNA为模板合成另一条与其互补的单链DNA,两条互补的单链DNA分子组成一个双链cDNA分子。因此,双链cDNA分子的序列同转录产生的mRNA分子的基因是相同的。所以一个cDNA分子就代表一个基因。但是cDNA仍不同于基因,因为基因在转录产生mRNA时,一些不编码的序列即内含子被删除了,保留的只是编码序列,即外显子。所以cDNA序列都比基因序列要短得多,因为cDNA中不包括基因的非编码序列---内含子。
技术发展
垃圾DNA
一项针对基因组进行的广泛比较研究显示,问题的答案可能就隐藏在生物的垃圾脱氧核糖核酸(DNA)中。美国科学家发现,生物越复杂,其携带的垃圾DNA就越多,而恰恰是这些没有编码的“无用”DNA帮助高等生物进化出了复杂的机体。
自从第一个真核生物——包括从酵母到人类的有细胞核的生物——的基因组被破译以来,科学家一直想知道,为什么生物的大多数DNA并没有形成有用的基因。从突变保护到染色体的结构支撑,对于这种所谓的垃圾DNA的可能解释有许多种。但是2004年从人类、小鼠和大鼠身上得到的完全一致的关于垃圾DNA的研究结果却表明,在这一区域中可能包含有重要的调节机制,从而能够控制基础的生物化学反应和发育进程,这将帮助生物进化出更为复杂的机体。与简单的真核生物相比,复杂生物有更多的基因不会发生突变的事实无疑极大地强化了这一发现。
为了对这一问题有更深的了解,由美国加利福尼亚大学圣塔克鲁斯分校(UCSC)的计算生物学家David Haussler领导的一个研究小组,对5种脊椎动物——人、小鼠、大鼠、鸡和河豚——的垃圾DNA序列与4种昆虫、两种蠕虫和7种酵母的垃圾DNA序列进行了比较。研究人员从对比结果中得到了一个惊人的模式:生物越复杂,垃圾DNA似乎就越重要。
这其中暗含的可能性在于,如果不同种类的生物具有相同的DNA,那么这些DNA必定是用来解决一些关键性的问题的。酵母与脊椎动物共享了一定数量的DNA,毕竟它们都需要制造蛋白质,但是只有15%的共有DNA与基因无关。研究小组在2005年7月14日的《基因组研究》杂志网络版上报告说,他们将酵母与更为复杂的蠕虫进行了比较,后者是一种多细胞生物,发现有40%的共有DNA没有被编码。随后,研究人员又将脊椎动物与昆虫进行了对比,这些生物比蠕虫更为复杂,结果发现,有超过66%的共有DNA包含有没有编码的DNA。
参与该项研究工作的UCSC计算生物学家Adam Siepel指出,有关蠕虫的研究结果需要慎重对待,这是由于科学家仅仅对其中的两个基因组进行了分析。尽管如此,Siepel还是认为,这一发现有力地支持了这样一种理论,即脊椎动物和昆虫的生物复杂性的增加主要是由于基因调节的精细模式。
DNA探针
DNA探针是最常用的核酸探针,指长度在几百碱基对以上的双链DNA或单链DNA探针。现已获得DNA探针数量很多,有细菌、病毒、原虫、真菌、动物和人类细胞DNA探针。这类探针多为某一基因的全部或部分序列,或某一非编码序列。这些DNA片段须是特异的,如细菌的毒力因子基因探针和人类Alu探针。这些DNA探针的获得有赖于分子克隆技术的发展和应用。以细菌为例,加之分子杂交技术的高敏感性,分子杂交在临床微生物诊断上具有广阔的前景。
细菌的基因组大小约5×106bp,约含3000个基因。各种细菌之间绝大部分DNA是相同的,要获得某细菌特异的核酸探针,通常要采取建立细菌基因组DNA文库的办法,即将细菌DNA切成小片段后分别克隆得到包含基因组的全信息的克隆库。然后用多种其它菌种的DNA作探针来筛选,产生杂交信号的克隆被剔除,最后剩下的不与任何其它细菌杂交的克隆则可能含有该细菌特异性DNA片段。将此重组质粒标记后作探针进一步鉴定,亦可经DNA序列分析鉴定其基因来源和功能。因此要得到一种特异性DNA探针,常常是比较繁琐的。探针DNA克隆的筛选也可采用血清学方法,所不同的是所建DNA文库为可表达性,克隆菌落或噬斑经裂解后释放出表达抗原,然后用来源细菌的多克隆抗血清筛选阳性克隆,所得到多个阳性克隆再经其它细菌的抗血清筛选,最后只与本细菌抗血清反应的表达克隆即含有此细菌的特异性基因片段,它所编码的蛋白是该菌种所特有的。用这种表达文库筛选得到的显然只是特定基因探针。
DNA修复
DNA修复(DNA repairing)是细胞对DNA受损伤后的一种反应,这种反应可能使DNA结构恢复原样,重新能执行它原来的功能;但有时并非能完全消除DNA的损伤,只是使细胞能够耐受这DNA的损伤而能继续生存。也许这未能完全修复而存留下来的损伤会在适合的条件下显示出来(如细胞的癌变等),但如果细胞不具备这修复功能,就无法对付经常在发生的DNA损伤事件,就不能生存。
DNA复制
DNA复制是指DNA双链在细胞分裂以前进行的复制过程,复制的结果是一条双链变成两条一样的双链(如果复制过程正常的话),每条双链都与原来的双链一样。这个过程是通过名为半保留复制的机制来得以顺利完成的。复制可以分为以下几个阶段:
DNA片段的生成:在引物提供了3'-OH末端的基础上,DNA聚合酶催化DNA的两条链同时进行复制过程,由于复制过程只能由5'->3'方向合成,因此一条链能够连续合成,另一条链分段合成,其中每一段短链成为冈崎片段(Okazaki fragments)。
RNA引物的水解:当DNA合成一定长度后,DNA聚合酶水解RNA引物,补填缺口。
DNA连接酶将DNA片段连接起来,形成完整的DNA分子。
最后DNA新合成的片段在旋转酶的帮助下重新形成螺旋状。
PCR复制技术

脱氧核糖核酸
DNA重组
重组DNA是一种人工合成的脱氧核糖核酸。它是把一般不同时出现的DNA序列组合到一起而产生的。从遗传工程的观点来看重组DNA是把相关的DNA添加到已有生物的基因组中,比如细菌的质粒中,其目的是为了改变或者添加特别是的特性,比如免疫。重组DNA与遗传重组不是一回事。它不是重组细胞内或者染色体上已经存在的基因组,而完全是通过外部工程达到的。重组蛋白质是从重组DNA合成出来的蛋白质。
超速离心
近代质粒DNA分离纯化以从大肠杆菌中分离为代表,鉴于大肠杆菌(E.coli)在分子生物学研究中的重要地位,从大肠杆菌(E.coli)中分离纯化质粒DNA(Plasmid DNA)成为超离心技术中一个重要课题。而质粒DNA的快速分离纯化又对超离心设备(超速离心机、转头和附属设备)提出了更高要求。
质粒DNA超速离心的分离方法
传统的分离方法:数年前,由于受设备条件限制,质粒DNA的分离一般用CsCl平衡等密度离心法,自形成梯度。以10~12ml单管容量为例,用甩平转头分离,36.000rpm×60小时,用角式转头分离45,000rpm×36小时,前者包括加减速在内共用去1.3亿转驱动部寿命,后者也要用去1亿转驱动部寿命,这对当时超速离心机总寿命为100~200亿转来看,无疑每次实验费用过高,加上CsCl用量多、价格贵等因素,使这类分离纯化工作成为非常昂贵的实验。
质粒DNA超速离心分离的最新进展
1、超速垂直管转头的离心分离(钦合金或碳纤维制造的):从1975年垂直管转头向世后,最高转速从50,000rpm到120,000rpm,RCFmax可达700,000Xg,90年代开发的新机型和转头己能够使质粒DNA垂直管离心分离实验做起来得心应手。
2、近垂直管转头离心分离:为了消除垂直管转头用于质粒DNA离心在壁部形成的RNA沉淀对已形成的DNA区带的污染,同时也为了改进一般斜角式转头(倾角25——35)由于沉降距离较长,因而分离时间也较长的缺点,近几年开发了多种近垂直管转头(即Near VerticalTube Rot时,简称NVT转头或Neo Angle Rotor,小假角转头,简称NT).它们的离心管纵剖面中心轴线与离心机驱动轴线之间夹角在7.5——10之间,转速从65,000rpm到120,OOOrpm,RCFmax可达646,000×g单管容量从2ml至13.5ml。NVT(或NT)转头的开发主要是为质粒DNA分离而设计,当然它也适用于线粒体DNA、染色体DNA、RNA及血清脂蛋白的分离·纯化。
3、不连续阶梯梯度分离:质校DNA分离纯化传统方法是采用金管CsCl自形成梯度平衡等密度离心法,离心开始时金管CsCl密度均一,样品均匀分布其中。
应用领域
身份鉴定
一个人有23对(46条)染色体,同一对染色体同一位置上的一对基因称为等位基因,一般一个来自父亲,一个来自母亲。如果检测到某个DNA位点的等位基因,一个与母亲相同,另一个就应与父亲相同,否则就存在疑问了。
利用DNA进行亲子鉴定,只要作十几至几十个DNA位点作检测,如果全部一样,就可以确定亲子关系,如果有3个以上的位点不同,则可排除亲子关系,有一两个位点不同,则应考虑基因突变的可能,加做一些位点的检测进行辨别。DNA亲子鉴定,否定亲子关系的准确率几近100%,肯定亲子关系的准确率可达到99.99%。
DNA(脱氧核糖核酸)是人身体内细胞的原子物质。每个原子有46个染色体,另外,男性的精子细胞和女性的卵子,各有23个染色体,当精子和卵子结合的时候。这46个原子染色体就制造一个生命,因此,每人从生父处继承一半的分子物质,而另一半则从生母处获得。
DNA亲子鉴定测试与传统的血液测试有很大的不同。它可以在不同的样本上进行测试,包括血液,腮腔细胞,组织细胞样本和精液样本。由于血液型号,例如A型,B型,O型或RH型,在人口中比较普遍,用于分辨每一个人,便不如DNA亲子鉴定测试有效。除了真正双胞胎外,每人的DNA是独一无二的.。由于它是这样独特,就好像指纹一样,用于亲子鉴定,DNA是最为有效的方法。我们的结果通常是比法庭上要求的还准确10到100倍。
通过遗传标记的检验与分析来判断父母与子女是否亲生关系,称之为亲子试验或亲子鉴定。DNA是人体遗传的基本载体,人类的染色体是由DNA构成的,每个人体细胞有23对(46条)成对的染色体,其分别来自父亲和母亲。夫妻之间各自提供的23条染色体,在受精后相互配对,构成了23对(46条)孩子的染色体。如此循环往复构成生命的延续。
由于人体约有30亿个碱基对构成整个染色体系统,而且在生殖细胞形成前的互换和组合是随机的,所以世界上没有任何两个人具有完全相同的30亿个核苷酸的组成序列,这就是人的遗传多态性。尽管遗传多态性的存在,但每一个人的染色体必然也只能来自其父母,这就是DNA亲子鉴定的理论基础。
传统的血清方法能检测红细胞血型、白细胞血型、血清型和红细胞酶型等,这些遗传学标志为蛋白质(包括糖蛋白)或多肽,容易失活而导致检材得不到理想的检验结果。此外,这些遗传标志均为基因编码的产物,多态信息含量(PIC)有限,不能反映DNA编码区的多态性,且这些遗传标志存在生理性、病理性变异(如A型、O型血的人受大肠杆菌感染后,B抗原可能呈阳性。因此,其应用价值有限。
DNA检验可弥补血清学方法的不足,故受到了法医物证学工作者的高度关注,近几年来,人类基因组研究的进展日新月异,而分子生物学技术也不断完善,随着基因组研究向各学科的不断渗透,这些学科的进展达到了前所未有的高度。在法医学上,STR位点和单核苷酸(SNP)位点检测分别是第二代、第三代DNA分析技术的核心,是继RFLPs(限制性片段长度多态性)VNTRs(可变数量串联重复序列多态性)研究而发展起来的检测技术。作为最前沿的刑事生物技术,DNA分析为法医物证检验提供了科学、可靠和快捷的手段,使物证鉴定从个体排除过渡到了可以作同一认定的水平,DNA检验能直接认定犯罪、为凶杀案、强奸杀人案、碎尸案、强奸致孕案等重大疑难案件的侦破提供准确可靠的依据。随着DNA技术的发展和应用,DNA标志系统的检测将成为破案的重要手段和途径。此方法作为亲子鉴定已经是非常成熟的,也是国际上公认的最好的一种方法。特别提到一点:同卵双胞胎的DNA检测结果是一样的。
在网上发布的遗传数据,那些来自1000多人的长达几十亿个DNA字母的串子,看似是完全匿名的。但仅仅靠一些网上的聪明侦探手段,一位遗传学研究者就把从研究对象组中随机选出的5个人的身份确定了出来。不仅如此,他还找到他们的整个家族,确定了近50个人的身份,虽然这些亲属与研究一点也不沾边。
这位研究者并未公布他所发现的人的姓名,但这项发表在周四的《科学》(Science)杂志上的工作表明,保护参加医学研究的志愿者的隐私不是一个简单的事情,因为他们提供的遗传信息需要公开,以便科学家使用。
研究人员表示,“让认为能够完全保护隐私或使数据匿名的幻想继续下去,已不再是一个可维持的立场。”
应用案例
2007年,迷恋研究此案的爱德华兹在一次拍卖会上买下一条带有血迹的披肩,据称为妓女凯瑟琳·埃多斯凶杀案现场物品。
2014年9月7日,英国商人拉塞尔·爱德华兹和法医学专家,借助先进的法医分析技术,成功破解困扰世人126年的谜:谁是英国连环杀手“开膛手杰克”。借助分析和比对DNA样本,认定波兰美发师阿伦·科斯明斯基为真凶。
科斯明斯基是犹太人,他被警方列为3名重点嫌疑人之一,一名目击者也指认他为凶手。但是,警方没有足够证据指控科斯明斯基。他最终于53岁时死在精神病院。
爱德华兹锁定了科斯明斯基。基因证据专家采用“真空吸取”的方式获取了DNA样本,与埃多斯后裔的DNA比对后,确定披肩上的血迹属于埃多斯。
发展计划
人类基因组计划(human genome project,HGP)是由美国科学家于1985年率先提出,于1990年正式启动的。美国、英国、法兰西共和国、德意志联邦共和国、日本国和中国科学家共同参与了这一价值达30亿美元的人类基因组计划。这一计划旨在为30多亿个碱基对构成的人类基因组精确测序,发现所有人类基因并确定其在染色体上的位置,破译人类全部遗传信息。与曼哈顿原子弹计划和阿波罗登月计划并称为三大科学计划。
2000年6月26日,参加人类基因组工程项目的美国、英国、法国、德国、日本和中国,六国科学家共同宣布,人类基因组草图的绘制工作已经完成。最终完成图要求测序所用的克隆能忠实地代表常染色体的基因组结构,序列错误率低于万分之一。95%常染色质区域被测序,每个Gap小于150kb。完成图将于2003年完成,比预计提前2年。
在人体全部22对常染色体中,1号染色体包含基因数量最多,达3141个,是平均水平的两倍,共有超过2.23亿个碱基对,破译难度也最大。一个由150名英国和美国科学家组成的团队历时10年,才完成了1号染色体的测序工作。
科学家不止一次宣布人类基因组计划完工,但推出的均不是全本,这一次杀青的“生命之书”更为精确,覆盖了人类基因组的99.99%。解读人体基因密码的“生命之书”宣告完成,历时16年的人类基因组计划书写完了最后一个章节。
作为人类基因组计划的后续计划,The ENCODE Project 在2003年9月启动的跨国研究项目。该项目旨在解析人类基因组中的所有功能性元件。该项目联合了来自美国,英国,西班牙,新加坡和日本的32个实验室的422名科学家的努力,获得了迄今最详细的人类基因组分析数据(他们获得并分析了超过15兆兆字节的原始数据)。研究花费了约300年的计算机时间,对147个组织类型进行了分析,以确定哪些能打开和关闭特定的基因,以及不同类型细胞之间的“开关”存在什么差异。
2012年9月5日,ENCODE项目的阶段性研究结果被整理成30篇论文发表于《自然》(6篇),《基因组研究》(6篇)和《基因组生物学》(18篇)上。研究结果显示,人类基因组内的非编码DNA至少80%是有生物活性的,而并非之前认为的“垃圾” DNA (junk DNA)。这些新的发现有望帮助研究人员理解基因受到控制的途径,以及澄清某些疾病的遗传学风险因子。
2012年12月21日,ENCODE项目被《科学》杂志评为本年度十大科学突破之一。
2012年12月28日,早老素同源蛋白PSH的晶体结构。
生物功能
在基因组中,遗传信息存储在称为基因的DNA序列中,这个遗传信息的传递由互补的含氮碱基序列的存在得到保证。事实上,在转录过程中,遗传信息可以很容易地被转录到互补的RNA链中(mRNA)。mRNA通过翻译合成蛋白质。或者,细胞可以通过称为DNA复制的过程简单地复制遗传信息。
基因组结构
真核生物基因组DNA位于细胞核内,线粒体和叶绿体内也有DNA。原核生物DNA被包裹在细胞质中不含细胞膜的不规则细胞器类核中。遗传信息包含在基因中,基因是能够影响生物体表型的遗传单位。每个基因含有开放阅读框(能够转录成RNA的区域)和由启动子和增强子组成的调节区。在许多物种中,只有一小部分基因组序列可以被转录和翻译。例如,人类基因组中只有1.5%序列含有编码蛋白质的外显子,超过50%的人类基因组由重复的非编码DNA序列组成。在任何情况下,不编码蛋白质的DNA序列也可以转录成非编码RNA,参与基因表达的调控。一些非编码序列是对染色体的结构组成部分。端粒和着丝粒区域通常含有非常少的基因,但对于染色体的功能和稳定性是必需的。
转录和翻译
基因是含有能够影响生物体表型特征的遗传信息的DNA序列。基因内的DNA碱基序列作为模板可以合成RNA分子,在大多数情况下,RNA分子被翻译成多肽,最终称为蛋白质。将基因的核苷酸序列复制到RNA链中的过程称为转录,由RNA聚合酶催化发生。 RNA链有不同的命运:一些RNA分子实际上具有结构(例如在核糖体内发现的那些rRNA)或催化(如核酶)功能;绝大多数RNA经历成熟过程产生mRNA,被翻译成蛋白质。翻译过程发生在细胞质中,其中mRNA与核糖体结合,并由遗传密码介导。核糖体允许顺序读取mRNA密码子,有利于它们识别和与特定tRNA相互作用,这些tRNA携带对应于每个单个密码子的氨基酸分子。
相互作用
所有DNA功能都取决于其与特定蛋白质的相互作用。这些相互作用可以是非特异性的,也可以是极其特异性的。还有许多可以结合DNA的酶,其中,在DNA转录和复制中复制DNA序列的聚合酶特别重要。
结合DNA蛋白质
结构蛋白可与DNA结合,是非专一性DNA-蛋白质交互作用的常见例子。染色体中的结构蛋白与DNA组合成复合物,使DNA组织成紧密结实的染色质构造。对真核生物来说,染色质是由脱DNA与一种称为组织蛋白的小型碱性蛋白质所组合而成;而原核生物体内的此种结构,则掺杂了多种类型的蛋白质。
DNA可在组织蛋白的表面上附着并缠绕整整两圈,以形成一种称为核小体的盘状复合物。组织蛋白里的碱性残基,与DNA的酸性糖磷酸骨架之间可形成离子键,使两者发生非专一性交互作用,也使复合物中的碱基序列相互分离。
在碱性氨基酸残基上所发生的化学修饰有甲基化、磷酸化与乙酰化等,这些化学作用可使DNA与组织蛋白之间的作用强度发生变化,进而使DNA与转录因子接触的难易度改变,影响转录作用的速率。其他位于染色体内的非专一性DNA结合蛋白,还包括一种能优先与DNA结合,并使其扭曲的高移动性群蛋白。这类蛋白质可以改变核小体的排列方式,产生更复杂的染色质结构。
DNA结合蛋白中有一种专门与单链DNA结合的类型,称为单链DNA结合蛋白。人类的复制蛋白A是此类蛋白中获得较多研究的成员,作用于多数与解开双螺旋有关的过程,包括DNA复制、重组以及DNA修复。这类结合蛋白可固定单链DNA,使其变得较为稳定,以避免形成茎环(stem-loop),或是因为核酸酶的作用而水解。
相对而言,其他的蛋白质则只能与特定的DNA序列进行专一性结合。大多数关于此类蛋白质的研究集中于各种可调控转录作用的转录因子。这类蛋白质中的每一种,都能与特定的DNA序列结合,进而活化或抑制位于启动子附近序列的基因转录作用。转录因子有两种作用方式,第一种可以直接或经由其他中介蛋白质的作用,而与负责转录的RNA聚合酶结合,再使聚合酶与启动子结合,并开启转录作用。第二种则与专门修饰组织蛋白的酵素结合于启动子上,使DNA模板与聚合酶发生接触的难度改变。
由于目标DNA可能散布在生物体中的整个基因组中,因此改变一种转录因子的活性可能会影响许多基因的运作。这些转录因子也因此经常成为信号传递过程中的作用目标,也就是作为细胞反映环境改变,或是进行分化和发育时的媒介。具专一性的转录因子会与DNA发生交互作用,使DNA碱基的周围产生许多接触点,让其他蛋白质得以“读取”这些DNA序列。多数的碱基交互作用发生在大凹槽,也就是最容易从外界接触碱基的部位。
DNA
核酸酶和连接酶:核酸酶是能够切割DNA链的酶,因为它们催化磷酸二酯键的水解。从位于DNA链末端的核苷酸开始水解DNA的核酸酶称为核酸外切酶。另一方面,直接切入DNA链的那些是内切核酸酶。分子生物学中使用最广泛的核酸酶,称为限制性内切酶,以切割特定序列的DNA。在自然界中,这种酶通过在进入细菌细胞时消化噬菌体DNA来保护细菌免受噬菌体感染。通常,限制性核酸酶识别特定的回文核苷酸序列,称为限制性位点。这些酶广泛用于涉及在载体内亚克隆DNA的技术中。
DNA连接酶:是能够使用来自ATP或NAD的化学能将先前切割或断裂的DNA链聚集在一起的酶。连接酶在DNA滞后链复制中特别重要,因为它们将冈崎碎片组合成DNA链。连接酶在DNA修复和基因重组中也发挥重要作用。
拓扑异构酶和解旋酶:拓扑异构酶是具有活性核酸酶和连接酶的酶。这些酶能够改变DNA的拓扑特性。它们中的一些通过切割DNA螺旋并允许其旋转,降低其超螺旋程度,然后通过连接酶将两端连接。另一方面,其它拓扑异构酶能够在连接断裂的DNA链之前,切断螺旋,并允许第二个螺旋通过断裂部位。拓扑异构酶是许多涉及DNA的过程所必需的,例如DNA复制和转录。螺旋酶是能够利用核苷三磷酸中存在的化学能的蛋白质,尤其是ATP,以破坏核碱基之间形成的氢键,从而允许DNA的双螺旋打开成单链。
聚合酶:聚合酶是从核苷三磷酸合成多核苷酸链的酶。它们通过向链上存在的先前核苷酸的3'-OH添加核苷酸起作用。因此,所有聚合酶都以5' - 3'方向起作用。DNA复制需要DNA依赖的DNA聚合酶,实现DNA序列的完美拷贝。有些DNA聚合酶具有校对功能,能够检测含氮碱基之间的错配错误并激活3'或5'外切核酸酶作用以去除不正确的碱基。在大多数生物体中,DNA聚合酶在称为replisoma的较大蛋白质复合物中起作用,该复合体由许多酶例如解旋酶组成。 RNA依赖的DNA聚合酶是使用RNA片段作为模板合成DNA的特殊类聚合酶,包括逆转录酶(一种参与逆转录病毒感染的病毒酶)和端粒酶(它是端粒复制所必需的)。与DNA依赖性DNA聚合酶一样,这些RNA依赖的DNA聚合酶也在由辅助分子和调节分子组成的广泛蛋白质复合物中起作用。
有关报道
2022年12月14日,据新民晚报消息,《自然》(《Nature》)杂志发布的最新封面研究论文显示,研究团队分离出200万年前的环境DNA片段,而曾经发现的最古老DNA序列时间是120万年,几乎翻倍。
参考资料
[1]
警惕!日本病毒学家:英国发现的变异株毒力更强2021-01-23T15:22:22+08:00[引用日期2022-06-13 19:12:55]
[2]
人类基因组工作草图绘制完成2021-06-27T10:01:13+08:00[引用日期2022-06-13 19:13:08]
[3]
当基因可以编辑,可以为人类带来什么?2020-09-22T20:46:32+08:00[引用日期2022-06-14 10:17:09]
[4]
外媒:人类“完整说明书”出炉 全部基因组测序完成2021-06-17T10:25:14+08:00[引用日期2022-06-14 11:01:48]
[5]
碱基精度99.9999% 高质量拟南芥基因组完成组装2021-09-15T09:46:53+08:00[引用日期2022-06-14 11:02:10]
展开
相关合集

生物分子结构参阅
共6个词条1.2w阅读

DNA双螺旋
核酸的构象之一
核糖核酸
遗传信息载体
核酸
一种由许多核苷酸聚合成的化合物
查看更多
相关视频
全部
独家
1201次播放01:23
生物遗传的关键物质——脱氧核糖核酸
简介
1832次播放02:34
生命的蓝图 DNA的发现与探索
更多知识点


