进化(生物学名词)

VLoG
次浏览
更新时间:2023-05-18
进化
本词条是多义词,共13个义项
生物学名词

基本信息
| 中文名 | 进化 |
| 别名 | 演化 |
| 外文名 | evolution |
| 释义 | 生物种群在世代的遗传变化 |
| 反义词 | 退化 |
展开
进化介绍

人类进化
达尔文并未对“evolution”下过定义,在1859年出版的《物种起源》第一版中,也并未使用“evolution”这个字,当时达尔文是使用“经过改变的继承”(descent with modification)、“改变过程”(processof modification)或是“物种改变的原理”(doctrine of the modification of species)等词。evolution这个字在当时生物学上的意义,是胚胎发育过程,在当时一般用语中具有“进步”的含义,而达尔文反对将“进步”之类的用语来描述生物改变的过程。他曾在《物种起源》第7章中说:“(天择)的最后结果,包括了生物体的进步(advance)及退步(retrogression)两种现象”。而后来包括达尔文在内的学者,之所以改用evolution来描述生物演化现象,乃因英国哲学家赫伯特·史宾赛在许多著作里进行的名词统一。
以自然选择为基础的进化理论,最早是由查尔斯·达尔文与亚尔佛德·罗素·华莱士所提出,详细论述则出现在达尔文出版于1859年的《物种起源》。1930年代,达尔文自然选择说与孟德尔遗传合二为一,形成了现代综合进化理论,连结了进化的“机制”(自然选择)与进化的“单位”(基因)。这种有力的解释及具预测性的理论,成为现代生物学的中心原则,使地球生物的多样性得以统一说明。

转座子的坟场——人类基因组
进化与演化
概况
最初中文对“evolution”这个字有两种翻译。“进化”一词来自日语(日制汉语)。严复是最早反对使用“进化”者之一。后人在《天演论》书尾的名词表中写到:“evolution一词,严氏译为天演,近人撰述多以进化二字当之。严复主张以'天演'取代'进化'。”

《天演论》
严复所翻译的不是《物种起源》,而是赫胥黎的《进化论与伦理》,多部分内容与生物学无关。赫胥黎于该书导言中设有一节,立evolution之界说,“谓为初指进化而言,继则兼包退化之义。严氏于此节略而未译,然其用天演两字,固守赫氏之说也”。也就是说,严复理解到单用“进化”一词的缺憾,故自创“天演”二字取代“进化”。早在达尔文逝世不久时,中文上用“进化论”之名是一致的,例如孙中山在著作中盛赞该论。除了台湾,“进化论”一词沿用至今。中国台湾地区的学者、政府机构、教科书等也一向沿用“进化论”,台湾地区教育事务主管部门却更改课本,以演化论代替,新旧学生被概念混淆,不知其是否为同样理论或是两套理论。
根据台湾地区教育事务主管部门所编辑的辞典,“进化”定义为生物由低级到高级、由简单到复杂的发展过程,并将“退化”定义为进化的反义词。演化则被定义为生物物种为了因应时空的嬗变,在形态和行为上与远祖有所差异的现象,演化明显较适用于生物学的讨论。
在翻译来说,主要考量的是达尔文所取英语原名的意义,由简单到复杂生物的出现确是有“进”的含义。而与达尔文自然选择论相对抗的如“智慧设计”论等,则否认眼睛这样精确和复杂的器官可以进化出来。
演化模型
微观演化与宏观演化
演化可以依据时间长短与差异程度,分成“微观演化”(微演化)与“宏观演化”(广演化)。微观演化指几个世代中,基因频率小范围的变化,例如现今世界各地人类的差异。宏观演化指长时间的演化过程,例如人类与灭绝祖先的关系。宏观演化的历史,包括生物群在化石纪录中突然出现突然丢失的环节、物种长期停滞不变等难以解释的现象,例如延续时间5370万年左右的寒武纪地层中保存大量化石的现象。
演化速率

红原鸡的演变
极端的渐变论认为,演化过程是以等速进行的,对于化石数量不平均问题,以“化石纪录本身并不完整”来解释。极端的渐变论,至今仍然经常使用在分子层次,并发展出分子时钟的观念。研究发现,分子时钟也并非等速变化,而是在不同物种或是不同年代具有不同的演化速度。不同的研究中,对分子演化速率的推算差异相当大。
进化证据
古生物学与化石纪录
古生物学是以生物化石为基础,以研究生物亲缘关系的一种研究。当生物个体死亡之后,它的尸体通常经由微生物分解而腐化,使得生存痕迹消失。但有时候这些遗迹可能会因为某些因素而被保存。只要是来自古代生物造成的痕迹,或是生物体本身,都可以称为化石。化石对于了解生物演化历程而言相当重要,因为它是较为直接的证据,且带有许多详细的资讯。在化石形成过程中,生物体的痕迹由于快速地受到掩埋,因此不会发生风化与分解的情形。较为常见的化石,通常源自骨骼或外壳等坚硬部位,并经由类似铸模的过程形成。坚硬的骨骼在动物死亡之后,会因为有机物的腐败,而产生一些漏洞。将骨骼掩埋的砂石或矿物,则会经由这些漏洞侵入骨骼内部,并将其填满。这种过程称为置换作用,属于型体的保留,而不是生物体本身的保留。也有一些化石是生物体本身,例如被冰冻的猛犸象、琥珀里的昆虫。此外,古代动物的脚印、或植物在地底下因温度与压力作用而碳化,都可称为化石。
不同时代的生物化石,会出现的在不同的地层中,如此便能够研究古生物之间,以及它们与现代生物之间的关系。“失落的环结”指演化过程可能出现过,却尚未发现的物种。连接两个物种之间的化石,则称为“过渡化石”。例如可能位于鸟类与恐龙中间的始祖鸟(Archaeopteryx)化石,以及一种具有四肢的大型浅水鱼(Tiktaalik)可能是鱼类与两栖类的过渡化石。
化石纪录对于古生物的研究有所限制,因为形成化石并不容易。举例而言,软体动物身上并没有太多能够形成化石的部分,还有一些生物生存在难以形成化石的环境当中。即使化石形成之后,也有可能因为某些原因被摧毁,使得大多数化石皆是零散的状态,只有少数化石能够保持完整。而当演化上的改变在族群中只占有少部分,或是环境变化使族群规模缩小时,都会使它们形成化石的机率相对较小。此外,化石几乎无法用来研究生物内部器官构造和机制。
寒武纪大爆发是达尔文以来困惑科学界一个多世纪的科学难题。澄江动物群代表包括脊索动物在内的许多现生动物门和亚门的重要历史起点,为揭示寒武纪大爆发的性质和特征,见证高级分类单位起源在先、低级分类单位起源在后的“自上而下”的宏观演化模型,以及为探索动物(特别是脊索动物和节肢动物))起源等提供了重要证据,充实了演化论内涵。
最古老脊索动物化石的发现与研究首次再现了包括人类在内的脊椎动物的生命蓝本。这些化石(特别是海口虫)保存的完整性令人叹为观止,包括了脑、神经索、血管等软组织构造,既包含有脊椎动物特征,又具有相似文昌鱼的特征,为无脊椎动物与脊椎动物间建起了桥梁。
生物地理学与物种分布
A图显示古代大陆间有陆桥连结,因此动物可在大陆间自由迁徙。B图显示陆桥消失之后,各地物种受到孤立,并形成新物种。
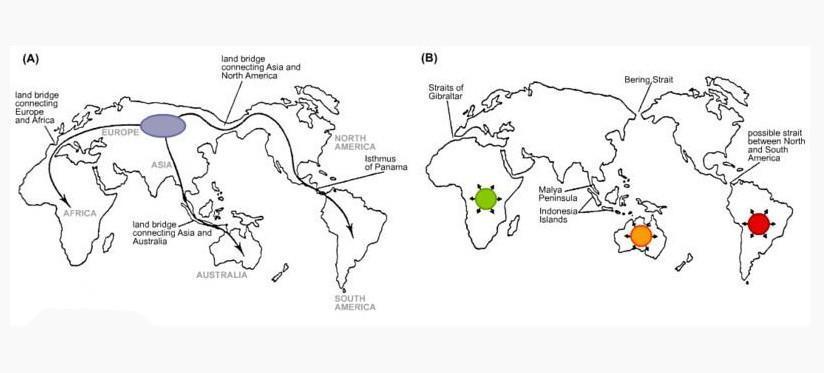
生物地理(左A图右B图)
除不同大陆之间有这种现象,大陆与其邻近岛屿因曾在地理上相连,因而也能够找到相似但变异了的物种。例如中国台湾、中国大陆与日本的猕猴之间的差异。
型态比较
对脊椎动物五趾肢的比较,支持了脊椎动物具有共同祖先的理论。举例而言,虽然人类、猫、鲸鱼与蝙蝠的五趾肢在型态上有所差异,但是主要架构都很相似。这些“同源”的构造,适应了不同的功能,如抓握、行走、游泳与飞行。
此外有一些构造在功能上相似,但却具有不同的型态。例如蝙蝠、鸟类与昆虫的翅膀;昆虫与脊椎动物的腿;章鱼与脊椎动物的眼睛;鱼类、鲸鱼与龙虾的鳍等。这类“异源”的构造,适应了相同的功能,如飞行、行走、感光与游泳。
发育过程
在发育初期都非常相似,在发育的过程中,这样的相似会逐渐减少,最后形成各物种的型态。举例而言,虽然各种成熟的脊椎动物差异很大,但是它们的胚胎型态在发育初期却非常相似,腮裂仍然出现在已经没有腮的爬虫类、鸟类与哺乳类胚胎中。鱼类的二房心会被保留到成体,而人类的胚胎虽然也有这种构造,却会在胚胎成熟后消失。

胚胎
在胚胎重演论提出直到被推翻的期间,胚胎学对于演化机制的解释并没有太大的进展。但是演化发育生物学(Evolitionary developmentalbiology,EvoDevo)研究,将分子生物学与发育生物学等学科结合,解释基因的改变对于动物形态的控制过程。同时也发现外表差异相当大的动物之间,也拥有相同的调控基因。以及相同的基因在不同的时间与空间,具有不同的作用。这些调控动物发育过程的基因,主要为一类Hox基因。
分子证据
分子生物学与生物化学可以利用构成生命的各种分子,来研究生物的亲缘关系。这些分子包括了核糖核酸(RNA)、脱氧核糖核酸(DNA)、蛋白质与糖体等。科学家已经可以用机器与电脑将这些分子的碱基序列、氨基酸序列定序。研究这些序列的学科分别为基因体学、蛋白质体学。此外还有一种更为复杂的糖体学,专门分析糖类构造。
基因体学透过脱氧核糖核酸序列的比较,可以建立出与传统分类学大致符合的种系发生树。这种方法通常用来确认并增强分类学的研究,将其它演化证据的漏洞修补或是修正。例如人类的DNA序列与黑猩猩的差异约为1.2%,与大猩猩的差异约为1.6%,与狒狒则为6.6%,如此就能推断并量化人类与猿类的亲缘关系。此外一种组成核糖体的16SrRNA序列,也是研究亲缘关系的重要分子,这些研究导致了三域系统的出现,将真核生物、古生菌与细菌分开。
蛋白质体学也支持了生物具有共同祖先的说法,因为许多生命所需的蛋白质,例如核糖体、DNA聚合酶与RNA聚合酶,不但出现在较原始的细菌中,也出在复杂的哺乳类体内。这些蛋白质的核心部分在不同生物中保有相似的构造与功能,而较复杂的生物具有较多的蛋白质次单位(protein subunit),以调控更复杂的蛋白质相互作用。
生物体内有一种脱氧核糖核酸片段,虽然与已知基因具有相似的序列,但是却无法发挥作用并完成蛋白质的制造。这类无用的核酸片段称为伪基因,是支持生物具有共同祖先的证据之一。它们之所以无法正常作用,是因为在演化过程中,基因突变累积过多的缘故。
抗药性
微生物与病毒的抗药性,在医学上是重要问题,也是演化证据之一。例如金黄葡萄球菌在1943年时仍可使用青霉素(盘尼西林)治疗,到了1947年就已经发现具抗药性的菌株。1960年代改用甲氧苯青霉素,同样因为抗药性菌种的散布,使得1980年代改用万古霉素,2002年时,已发现抗万古霉素的菌种。
遗传
进化中的变异
突变是指遗传的物质发生改变,广义的突变包括染色体数目和结构变异。不过一般所说的突变,是核酸序列的改变,也就是基因突变。基因突变是产生遗传变异的最根本原因,细胞中的遗传物质(通常是脱氧核糖核酸或核糖核酸)能够经由许多方式改变,例如细胞分裂时的复制错误、放射线的照射、化学物质的影响或是病毒感染。多细胞生物的基因突变,可依照发生的细胞种类分为两种。生殖细胞突变能够遗传到下一代,体细胞突变则通常限制在个体中。
基因突变可能对个体有害,也可能对个体有益,或是两者兼具。有害的隐性基因因为不会出现症状而被保留,当这些隐性基因配成对时,就可能使个体得到病变或是死亡。有一些基因虽然可能会造成病变,但是也可以使个体具有某些优势,例如带有一个镰刀型红血球疾病基因的人,对疟疾更有抵抗力。
对生物个体无益也无害的突变称为中性突变,在族群中的出现频率主要受到突变机率影响。由于这些突变不影响个体的生存机会,因此大多数物种的基因组在没有天择的状况下,依然会有稳定数量的的中性突变不断发生。单一碱基对的变换称为点突变,当一个或多个碱基对插入或是删除时,通常会使基因失去作用。
转座子(transposon)是生物的基因组片段,并且在基因组的演化上扮演重要角色。它们能够移动并插入基因组中,或是取代原有的基因,产生演化上的变异和多样性。DNA复制也被认为每百万年间,会在动物的基因组中产生数十到数百的新基因。
重组
在无性生殖的过程中,染色体上的任何一对等位基因都会遗传到下一代。但是对于行有性生殖的物种而言,亲代同源染色体中的等位基因,在制造生殖细胞的减数分裂过程中,会发生基因重组。这是一种不同的脱氧核糖核酸段落断裂并重新组合的过程。
基因重组
此外有性生殖中的孟德尔遗传规则,能够使有害的突变被清除,有益的突变被保留。当一个等位基因无法进行基因重组时(例如孤立的Y染色体),则不具有清除有害突变的效果,其有害突变逐渐累积,使族群的有效族群大小(effective populationsize/Ne)缩减,这种现象称为 希尔—罗伯森效应(Hill-Robertsoneffect)。若是染色体逐渐退化,则称为缪勒氏齿轮(Muller'sratchet),这种现象比较容易出现在无性生殖的生物中。
进化机制
概述
遗传变异一方面经由生殖而传递到下一个世代(被称为垂直基因转移),另一方面也可以透过水平基因转移(horizontal gene transfer/HGT),在物种之内或是物种之间传递。尤其是细菌经常使用这种方式交换基因,研究发现可能有跨物种的水平基因转移存在。基因流(geneflow)则是指基因在生物个体之间转移。
基因型
基因型(遗传因子)是产生表现型(外在表现)的根本。而表现型本身也拥有表型可塑性(phenotypicplasticity),能够在基因型未改变的状况下有所变化,并且能够遗传到下一世代。除了基因本身的改变,染色体的重新排列虽然不能改变基因,但是能够产生生殖隔离,并使新物种形成。
一般来说,选择包括了“天择”(自然选择)与“性择”(性选择)。天择的主要原因是物种所居住环境的改变,包括物种之间关系的变化;性择则是物种在繁殖的需求下而产生的选择。而这些性择所留下的性状,可能会有害于个体本身的生存能力。各种选择的分类事实上并不明确,也有一些分类以天择表示所有选择作用,并分为生态选择(ecological selection)与性择。
基因流
基因流也称为迁移(migration),当族群之间并未受到地理或是文化上的阻碍时,基因变异会经由一些个体的迁移,使基因在不同族群间扩散,这样的情形称为基因流。恩斯特·麦尔认为基因流类似一种均质化(homogenising)的过程,因此能够抵销选择适应的作用。当基因流受到某种阻碍,例如染色体的数目或是地理的隔阂,便会产生生殖隔离,这是物种形成的条件之一。
族群中等位基因的自由移动,也受到族群结构的阻碍,例如族群的大小或是地理分布。虽然理想状态中族群的生殖对象完全自由且完全随机,但是现实世界中并非如此,因此地理上的亲近程度会对这些基因的移动造成庞大的影响。而且当迁移数量较少的时候,基因流对演化的影响也较低。
基因漂变指的是族群中等位基因频率在每一个世代之间的随机的变化。这种变化能够以数学表达,哈迪-温伯格平衡描述了理想状态情况下(不考虑天择等因素)的数学模型。在理想状态中,后代的等位基因频率将接近随机分布。当族群规模较大,基因漂变的机率会较低;当族群规模较小的时候,基因漂变的现象较为明显。
当一个少数族群从原先族群之中分离而出,且两者的基因频率有所不同,若分离而出的少数族群与原先族群的基因无法继续交流,则两者的基因频率将渐行渐远。这种现象称为奠基者效应。例如从德国迁移到美国宾夕法尼亚的阿米什人,起源大约仅有200人,且习惯族内通婚。这个族群的埃利伟氏综合症(Ellis-van Creveld syndrome)出现频率较其他族群高。
水平基因转移
以16SrRNA的基因序列所建立的种系发生树,将生物演化历程分为三域系统,包括细菌、古菌与真核生物。第一个提出这种分类的是卡尔·乌斯(Carl Woese)。由于水平基因转移的存在,使生物的亲源关系可能复杂许多。
水平基因转移(horizontalgenetransfer/HGT),是个体将遗传物质传递到其它非本身后代个体的过程。这种机制使遗传物质得以在无直系关系的个体之间产生基因流。
水平基因转移也可以经由抗原转移(antigenicshift)、基因重整(reassortment)与杂交反应(hybridisation)等现象观察。病毒能够透过转导作用(transduction)在物种间传递基因。细菌则能够与死亡的细菌合体、经由转形作用(transformation),以及与活细菌进行接合(conjugation),而获得新的基因。而新的基因则能够以质体的形式,加入宿主细菌的基因组中。杂交的现象在植物中最显着,此外已知还有10种以上的鸟类物种能够杂交。另外在哺乳动物与昆虫中,也有杂交的例子,只是通常杂交所生的后代不具有生殖能力。HGT也是细菌传递抗药性的方式之一,而且有些发现表明HGT是原核生物与真核生物的演化重要机制。
由于HGT的存在,使种系发生学更加复杂,也使早期物种的演化过程出现一种隐藏关系(metaphor)。遗传资讯在生殖作用之外,也能在物种之间传递。这使科学家必须在解释演化关系的时候,表达出物种的隐藏关系,并且将不同的演化历程组合起来。
自然选择

孔雀开屏
由于各种基因的变异,使同一个族群中,不同个体的生存方式和繁殖方式有所不同。当环境发生改变,便会产生天择作用。之所以称为天择,是因为这种选择并非如基因漂变或基因突变一样随机,当环境改变发生时,将只有某些带有特定特征的群体能够通过这些考验。天择有一些特例,如被视为与天择拥有相等地位的选择方式,其中包括性择、人择等等。
性择指某个体比其它个体拥有较高的繁殖机会,因此它们的基因会被保留,使后代继续保有相同的优势。人择指人类为了本身的生存或是喜好而对不同的基因变异进行筛选,通常发生在农业、畜牧业或是宠物的育种上。此外,优生学则是人类对人类所进行的筛选行为。不过人类事实上只是自然界的一部分,因此人择与并天择没有质的分别。
这些特殊的选择机制,导致生物产生的适应环境的有益特征,并非会在演化过程中一定出现或是被保留。代表性生物有只能吃尼龙的尼龙菌,再如,拥有更多的手指对人类的生活可能会更加方便,但是这种方便几乎不会增加任何繁殖机会。
自然选择能使有利于生存与繁殖的遗传性状变得更为普遍,并使有害的性状变得更稀有。这是因为带有较有利性状的个体,能将相同的性状转移到更多的后代。经过了许多世代之后,性状产生了连续、微小且随机的变化,自然选择则挑出了最适合所处环境的变异,使适应得以发生。遗传漂变使性状在种群中所占比例产生的一些随机变化,来自一些使个体能够成功繁殖的偶然因素。
生命起源
对于最初始的生命起源目前尚未明了。而生物学的进化研究,通常不包括这段初始过程,因为这段过程牵涉到太阳系与地球的形成。所以对于生命起源的研究,许多是来自物理学与化学。例如1952年的米勒-尤列实验中,米勒(Stanley Miller)与尤列(Harold Urey)以氨、甲烷、氢气、氰酸与水等分子,模拟地球的原始状态,并首次在实验室中制造出氨基酸。而作为氨基酸原料的有机分子,有可能是来自太空中,或是海底火山。
而关于包括遗传物质在内的有机分子进化过程,科学家一般认为核糖核酸比蛋白质与脱氧核糖核酸更早出现,之后出现类似反转录酶的蛋白质,最后才有脱氧核糖核酸,不过这些理论的证据并不多。脂肪酸的出现则构成了原始的细胞膜,之后经由内共生等过程,形成最早的单细胞生物。
生命诞生的最初,地点很可能是位于地底深处的有着高温高压的有机物团,很大的一团,像海洋一样,这些有机物,在高温高压下,最初可能只是基本分子。分子之间会发生化合。
有机分子之间的化合,则缓慢的吸收了环境的热量,环境热能转化成了有机分子之间的化学能。
有机小分子化合之后,有的会解体,有的则是稳定的。
解体的,在高温和热流中还可以参与新的化合。
而稳定的,则有机会,与其他稳定的,再化合。增长增大,再稳定。
再稳定之后,还会有机会重复再化合增长再稳定,再增长的过程。
就像生命一样,选代。
在这个过程中,不断的吸收来自环境的热能。
环境热能被化合分子大量吸收之后,温度逐渐缓慢下降。
环境温度的降低,有利于有机分子状态的稳定,这最稳定的有机分子,则有机会,化合成更大的分子团。
化合的速度可能也加快了。
吸收环境热能的速度也会加快。
环境温度在有机分子团演进的过程中,因热能被分子团大量吸收而不断降低。降低的环境温度又有利于稳定的分子团继续扩大 规模。
但分子团太巨大了,不可能完全稳定,薄弱之处会产生断裂,然后可以与其他小分子团重结合。经过不断的断裂与交换而重结合成的大分子团,有机会再次稳定下来,成为一个稳定的整体,这样就可以继续结合,继续增大。迭代。
终于有一天,分子团无论如何不能继续稳定的增大了,它不断的组合又断裂,而它裂解下的碎片,则组成在了一起。成为一个与它本身结构相同,或者说近乎相同,应该不是一个,可能是几乎充满整个有机物的地底海洋的。
它个的出现,使得最初的有机物大分子,有机会和它们结合在一起。组成连合大分子团。并稳定下来。迭代。大分子团有一天,也会到极限。
但是其他的大分子团也已经产生了。
它们之间互相互换。
大分子团虽不能成为稳定的更大的分子团,但它可以在互相之间的交换中,暂时变的更大,更大。
更大,更大,更复杂。之后又变的更大,更大,更复杂。迭代,再迭代。演进。
巨大的分子团,总是不稳定的,它们周期性的成长,崩溃。
而分子团的中心,则是稳定的。
它不断的在外围成长再崩溃的过程中,转移。
而外围,也在演进,变的越来越稳定。
最终形成了稳定的细胞核和外围的细胞质细胞膜。
多细胞生物在演进中,细胞越来越多,个体越来越大,越来越复杂。
动物植物在演进中最终诞生了。
而所有的生命,演进并不会停止。
进化时间表
1、前寒武纪
已知的化石纪录中,最早生命遗迹是出现在约38亿年前,原核单细胞生物则出现在33亿年前。到了22亿年前,才出现最早的真核单细胞生物,如蓝绿菌。6亿年前藻类与软体无脊椎动物出现。在此之前的年代称为前寒武纪。
2、古生代

三叶虫化石。活跃于2-5亿万年前。
3、中生代
4、新生代
演化思想史
早期理论
早在古希腊时代,类似演化的思想已经出现,例如阿那克西曼德认为人类祖先来自海中。到了18世纪与19世纪,就已经有许多关于生命起源来自共同祖先的观念。建立生物学的拉马克,是第一位为演化提出科学理论的科学家。而达尔文与华莱士所提出,以天择为主要机制的演化论,成为第一个具有说服力的解释。在达尔文发表著作并成名之后,有许多先前尚未发表,发展较少的类似理论被发现。此外,达尔文在当时还未知任何遗传机制,因此他无法解释为何不同世代具有不同特征。尤其当时流行子代的性状为两位亲代性状混合的概念,使得任何变异理论上似乎会逐渐消失。
孟德尔发现遗传性状的分离现象,解决了原本性状混合的难题,然而一开始却受到忽略。而且原本遗传学家并不接受达尔文的天择观念,而是以突变作为演化原动力。直至20世纪,数学家罗纳德?费雪、生物学家莱特(SewallWright)与霍尔登(J.B.S.Haldane)才建立了群体遗传学,并与演化论结合。再加上汤玛斯·摩根、特奥多修斯·多布然斯基、朱利安、赫胥黎、恩斯特·麦尔、乔治·辛普森、斯特宾斯(G.L.Stebbins)等人的研究,又称为新达尔文主义的现代综合理论在1920年到1940年代开始成形。
到了1960年代,许多生物学家开始以基因中心演化观点探讨演化过程。道金斯更认为,基因是唯一的天择单位。此外还有汉弥尔顿提出以利他行为为基础的亲属选择。与之同时,古尔德与艾崔奇对演化的速率重新诠释,提出疾变平衡论,认为生物的演化速度是长期的停滞与短暂的爆发所组合。之后演化生物学成形,并且发展出许多分支。
当今研究与应用
演化生物学是研究演化的主要学门,探讨物种的起源和改变,以及物种之间的亲缘关系。这些研究影响了传统的分类学,并导致系统分类学的出现。演化发育生物学(evo-devo)比较不同动物在发育过程中的变化,由此探讨它们之间的关系与演化过程。体质人类学专注于人类的起源与演化,并探讨人种的差异,又称为生物人类学。
为了更深入研究演化过程与机制的细节,许多相关的分支学门产生。例如生态遗传、人类演化、分子演化与种系发生。由于生物学是奠基在其它更基础的自然科学之上,因此数学、统计学、物理学与化学对于了解演化机制也相当重要。例如为基因流、基因漂变等现象提供数学模式的群体遗传学,研究在演化动力影响下,等位基因的分布和改变。
遗传算法则是应用演化与遗传的各种机制,并结合电脑的运算能力来解决许多问题。应用的层面包含工程、设计与通讯科技等。
社会演化

社会演化
1976年,道金斯在《自私的基因》一书中,认为人类的文化也能以演化解释。他根据基因(gene)这个词,将文化的演化单位称为迷因(meme、在中文也被音译为迷米),类似遗传因子的基因,迷因为文化的遗传因子,也经由复制(模仿)、变异与选择的过程而演化。
争议
理论与事实
有时候演化会被人们强调为理论、学说或假说,而非真实。例如美国前总统里根曾在1980年的一场竞选活动中表示:“演化是一个理论,只是一个科学理论,直到现在依然在科学界中受到挑战,并且尚未被科学界认为绝对正确。”这类说法强调,演化只是一个理论,所以并不是真实存在的事物。然而对科学家而言,理论并非与真实对立。真实是指经验上所得的资料或数据,理论则是对事实的解释与想法。
进步、复杂化与退化
有些物种(如人类),常被认为是比其它的物种更高级,甚至是演化的方向与目的所在。且认为演化的过程必定会使生物愈来愈复杂,或是进行与演化相反的退化。生物学家认为演化是没有方向的过程,也没有任何预先计划的目标。虽然在已知的演化过程中,确实具有逐渐复杂的现象,但是依然有许多物种保持在较简单的状态,如细菌。因此复杂性可能增加也可能减少,或是维持不变,结果取决于天择的机制。
物种形成
物种形成有时被认为是无法直接观察的现象,并得出演化是不科学的结论。但科学发现不仅经由可重复实验得来,均变说(uniformitarianism)亦使科学家得以用经验来推论事物的原因。此外物种形成的例子也出现在植物中,还有刺鱼(stickleback)外胚叶发育不全(ectodysplasin)的等位基因,被用来当作研究基因转变与物种形成的模型。
有一种类似的观点,认为微观演化是可以观察的,而宏观演化则无法观察。但是由于宏观演化的机制与微观演化相同,所以 宏观演化事实上已经在微观演化中被观察。而且物种之间基因序列的比较,也显示少量的遗传变异,就可以导致外表相当大的变化。
熵与生命
有些观点认为演化增加复杂性的情形,违反了热力学第二定律。该定律是指在一个孤立系统中,熵只会增加或是维持不变,可以使用的自由能逐渐减少,最终反应逐渐趋于动态平衡。熵即物理学上的无序度。这种观点忽略了生态系统事实上并非孤立系统,所有生态系中生物所获得的自由能,都是来自太空中,尤其是太阳。太阳、地球与太空的系统并不违反热力学第二定律,因为太阳与地球辐射所产生的自由能,远超过生物演化所需。
演化的结果
适应
在天择的作用影响之后,生物能够更加适应它们所处的环境。只要是能够使个体拥有更大生存优势的过程,都可以称为适应。不过需要注意的是,适应并非放诸四海皆准,在一个环境中拥有优势的特征,可能会在另一个环境中成为缺陷,这种现象也能解释为何演化并没有任何默认方向和目的,只有适应或是不适应。
即使“进步”在演化过程中并非必要,但是物种之间的竞争关系,仍能使物种在最基本的环境适应之外,进行更进一步的变化。这些竞争类似人类的军备竞赛(armsrace),依照物种的关系可将其分成两种。一种是不对称竞争,指物种受到不同条件的选择,如掠食者与猎物的关系;另一种是对称竞争,指物种受到相同的条件选择,如森林中树木对阳光的争取。此外性择也具有竞争的特性,有些物种在繁殖机会的竞争压力之下,会逐渐发展出相当奇特的外观或行为,例如鲜艳羽毛与吞食异性。
物种形成
物种是指一群可以互相进行繁殖行为的个体。当一个物种分离成其交配行为受阻碍的不同种群时,再加上突变、遗传漂变,不同环境对于不同性状的青睐,便会使变异逐代累积,进而产生新物种。生物之间的相似性显示,所有已知物种,皆是从共同祖先或是祖先基因池逐渐分化产生而来。例如野猪在全世界有27个亚种。亚种间和亚种内核型都有一些差异,染色体数(2n)在36-38之间,如西欧野猪2n=36或37,日本野猪2n=38,但彼此间没有繁殖障碍。普氏野马66个染色体,家马(64个染色体)可以杂交有可育的后代,。染色体数56的野牛和60对黄牛,58染色体的大额牛,杂交有可育后代。他们的基因排列都有很大的统一度。
物种形成受到许多类型的机制影响,主要可以分成两种类型。一种是 异域性物种形成(allopatricspeciation)。是发生在族群受到地理隔离,而形成新物种的情况。例如大峡谷两侧的松鼠,由于峡谷地形的阻隔,形成两个物种。不过地理隔离不一并会使物种分化,当两个物种再度接近,有可能会产生杂交。例如中国台湾地区的白头翁与乌头翁,原本受到山脉阻隔而发展出不同型态,纯种乌头翁却逐渐与白头翁交配而减少数量。一种是 同域性物种形成(sympatric speciation)。是指新物种与固有物种在相同的地区生存,因为繁殖或是沟通等行为而产生生殖隔离。例如印尼的一种蝙蝠由于声音的不同,产生三个从未杂交的族群。
另外还有两种较特殊的物种形成方式。边域性物种形成(peripatric speciation)介于异域性与同域性之间,指物种原先受地理隔离,之后因族群扩大而与固有物种接触,却已经形成两个物种。边域性物种形成是由恩斯特·麦尔所提出,这种理论同时支持了疾变平衡理论,且与奠基者效应有关,是小族群演化的例子。临域性物种形成(parapatric speciation)指物种虽然生存在相连的区域,但是因为交流的困难而产生新物种。最著名的现象是环状物种(ringspecies),例如北极圈周围的黑脊鸥(Larusargentatus)。
灭绝
灭绝指物种或是某个分类上的族群消失,并减少生物多样性。某一物种的最后个体死亡,就是物种灭绝的

最知名的已灭绝动物之一,渡渡鸟
名言
学术观点
参考资料
[1]
Emerman等复活的逆转录病毒名为Pan troglodytes内源性逆转录病毒 · 生物通[引用日期2014-04-18]
[2]
试与命运共进退 · 人民论坛网[引用日期2022-10-28]
相关视频
全部
2.6万次播放01:40
如果人类继续进化200万年,到底会长什么样?


