神经元细胞(神经元细胞)

VLoG
次浏览
更新时间:2023-05-23
神经元细胞
基本信息
| 中文名 | 神经元细胞 |
| 外文名 | Neurons |
| 别名 | 神经组织 |
| 结构组成 | 神经系统 结构和功能 |
说明
人的大脑不同于身体的任何器官,因为出生后,它还没有发育完全。它会在出生后的最初几个月或几年中,随着他的经历和外界刺激,继续发育。出生时人会有数以千亿计的脑细胞,当他们链接起来(也就是形成树突),才可以传导信息。如果这些神经元没有被链接,是会渐渐消失的。
所以,大脑的生长,大部分原因是树突数量和密度增加的结果。树突在大脑中是负责接受信息的角色,从出生到4岁前,树突的密集程度明显加大,树突越多说明接受信息的能力越强,人也就越聪明。
神经元分类
细胞结构
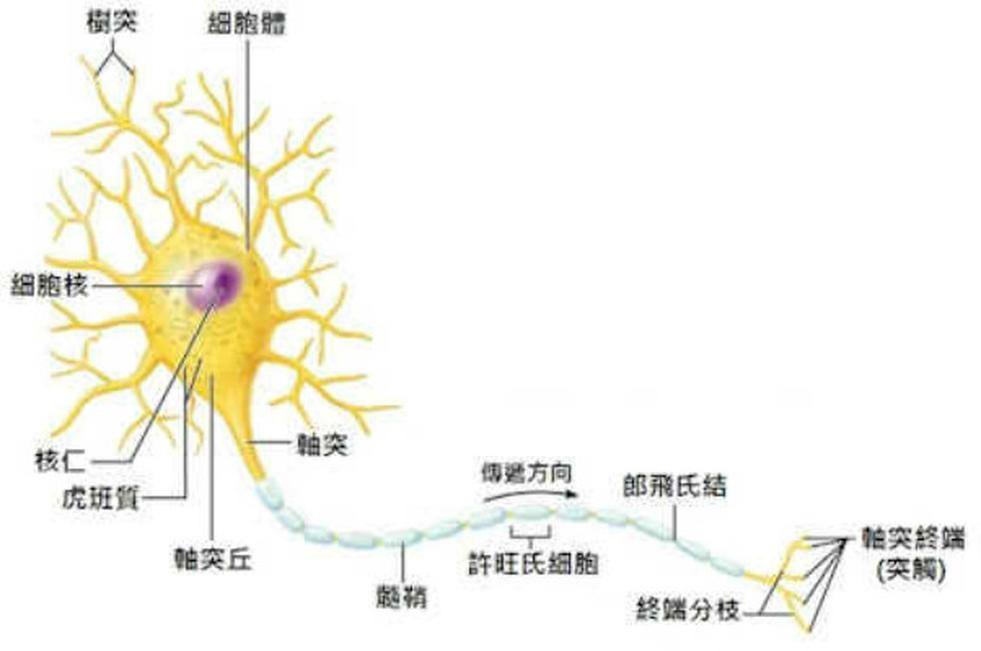
神经元的胞体(soma)在于脑和脊髓的灰质及神经节内,其形态各异,常见的形态为星形、锥体形、梨形和圆球形状等。胞体大小不一,直径在5~150μm之间。胞体是神经元的代谢和营养中心。
胞体的结构与一般细胞相似,有核仁、细胞膜、细胞质和细胞核。
(l)细胞膜:胞体的胞膜和突起表面的膜,是连续完整的细胞膜。除突触部位的胞膜有特优的结构外,大部分胞膜为单位膜结构。神经细胞膜的特点是一个敏感而易兴奋的膜。在膜上有各种受体(receptor)和离子通道(ionic chanel),二者各由不同的膜蛋白所构成。形成突触部分的细胞膜增厚。膜上受体可与相应的化学物质神经递质结合。当受体与乙酰胆碱递质或γ-氨基丁酸递质结合时,膜的离子通透性及膜内外电位差发生改变,胞膜产生相应的生理活动:兴奋或抑制。
(3)细胞质:位于核的周围,又称核周体(perikaryon)其中含有发达的高尔基复合体、滑面内质网,丰富的线粒体、尼氏体及神经原纤维,还含有溶酶体、脂褐素等结构。具有分泌功能的神经元,胞质内还含有分泌颗粒,如位于下丘脑的一些神经元。
l)尼氏体(Nissl body):又称嗜染质(chromophil substance),是胞质内的一种嗜碱性物质,在一般染色中岛被碱性染料所染色,多呈斑块状或颗粒状。它分布在核周体和树突内,而轴突起始段的轴丘和轴突内均无。依神经元的类型和不同生理状态,尼氏体的数量、形状和分布也有所差别。典型的如脊髓前角运动神经元,尼氏体数量最多,呈斑块状,分散于神经原纤维之间,有如虎皮样花斑,故又称虎斑小体(tigroid body)。而在脊神经节神经元的胞质内,尼氏体呈颗粒状,散在分布。电镜下,尼氏体是由许多发达的平行排列前粗面内质网及其间的游离核糖体组成。神经活动所儒的大量蛋白质主要在尼氏体合成,再流向核内、线粒体和高尔基复合体。当神经元损伤或中毒时,均能引起尼氏体减少,乃至消失。若损伤恢复除去有害因素后,尼氏体又可恢复。因此,尼氏体的形态结构可作为判定神经元功能状态的一种标志。
2)神经原纤维(neurofibril):在神经细胞质内,存在着直径约为2~3μm的丝状纤维结构,在银染的切片体本可清晰地显示出呈棕黑色的丝状结构,此即为神经原纤维,在核周体内交织成网,并向树突和轴突延伸,可达到突起的未消部位。在电镜下观察,神经原纤维是由神经丝甜神经微管集聚成束所构成。神经丝(neurofilament)或称神经细丝,是直径约为10nm细长的管状结构,是中间丝的一种,但与 其他细胞内的中间丝有所不同。在电镜高倍放大观察。可见神经细丝是极微细的管状结构,中间透明为管腔,管壁厚为3nm,其长度特长,多集聚成束。分散在胞质内,也延伸到神经元的突起中。神经丝的生理功能是参与神经元内的代谢产物和离子运输流动的通路。神经微管(neurotubule)是直径约25nm的细而长的圆形细管,管壁厚为5nm,可延伸到神经元的突起中,在胞质内与神经丝配列成束,交织成网。其生理功能主要参与胞质内的物质转运活动,接近微管表面的各种物质流速最大,微管的表面有动力蛋白(dynein),它本身具有ATP酶的作用,在ATP存在状态下,可使微管滑动,从而使微管具有运输功能。此外,还有较短而分散的微丝。微丝(microfilament)是最细的丝状结构,直径约5nm,长短不等,集聚成束,交织成网,广泛的分布在神经元的胞质和突起内,其主要功能具有收缩作用,适应神经元生理活动的形态改变。神经丝、微管、微丝,这三种纤维,构成神经元的细胞骨架(cytoskeleton),参与物质运输,在光镜下所显示仅是神经丝和神经微管形成的神经原纤维。 3)脂褐素(lipofuscin):常位于大型神经无核周体的一侧,呈棕黄色颗粒状,随年龄增长而增多,经电镜和组织化学证实为次级溶酶体形成的残余体(residual body),其内容物为溶酶体消化时残留的物质,多为异物、脂滴或退变的细胞器。某些神经元,如下丘脑,具有内分泌功能的分泌神经元(secretoryneuron),脑体内含直径IO0~30Onm的分泌颗粒,颗粒内含肽类激素(如加压素、催产素等)。
组成结构
神经元的突起是神经元胞体的延伸部分,由于形态结构和功能的不同,可分为树突和轴突。
树突
树突(dendrite)
是从胞体发出的一至多个突起,呈放射状。胞体起始部分较粗,经反复分支而变细,形如树枝状。树突的结构与脑体相似,胞质内含有尼氏体,线粒体和平行排列的神经原纤维等,但无高尔基复合体。在特殊银染标本上,树突表面可见许多棘状突起,长约0.5~1.0μm,粗约0.5~2.0μm,称树突棘(dendritic spine),是形成突触的部位。一般电镜下,树突棘内含有数个扁平的囊泡称棘器(spine apparatus)。树突的分支和树突棘可扩大神经元接受刺激的表面积。树突具有接受刺激并将冲动传入细胞体的功能。
轴突
轴突(axon)
每个神经元只有一根胞体发出轴突的细胞 质部位多呈贺锥形,称轴丘(axon hillock),其中没有尼氏体,主要有神经原纤维分布。轴突自胞体伸出后,开始的一段,称为起始段(initial segment),长约 15~25μm,通常较树突细,粗细均一,表面光滑,分支较少,无髓鞘包卷。离开胞体一定距离后,有髓鞘包卷,即为有髓神经纤维。轴突末端多呈纤细分支称轴突终未(axon terminal),与其他神经元或效应细胞接触。轴突表面的细胞膜,称轴膜(axolemma),轴突内的胞质称 轴质(axoplasm)或轴浆。轴质内有许多与轴突长袖平行的神经原纤维和细长的线粒体,但无尼氏体和高尔基复合体,因此,轴突内不能合成蛋白质。轴突成分代谢更新以及突触小泡内神经递质,均在胞体内合成,通过轴突内微管、神经丝流向轴突末端。神经元树突的末端可以接受其他神经传来的信号,并把信号传给神经元,因此是传入神经的末梢。而轴突的分枝可以把神经传给其他神经元或效应器,因此是传出神经的末梢。电镜下,从轴丘到轴突全长可见有许多纵向平行排列的神经丝和神经微管,以及连续纵行的长管状的滑面内质网和一些多泡体等。在高倍电镜下,还可见在神经丝、神经微管之间均有极微细纤维网络连接,这种横向连接的极细纤维称为微小梁(microtrabecula)起支持作用。轴突末端还有突触小泡。轴突运输(axonal transport)神经元的胞体和轴突在结构和功能上是一个整体,神经元代谢活动的物质多在胞体形成,神经元的整体生理活动物质代谢是由轴浆不断流动所实现。研究证明:神经元胞质自胞体向轴突远端流动,同时从轴突远端也向胞体流动。这种方向不同、快慢不一的轴质双向流动称为轴突运输。从胞体向轴突远端的运输,由于运输方向与轴质流动的方向一致故称为倾向运输(antrograde transport),这种运输有快慢之分:快速运输,其速度为每天200~500mm,是将神经元胞体合成的神经递质的各类小泡和有关的酶类等经长管状的滑面内质网和沿微管表面流向轴突末端,待神经冲动时释放。慢速运输也称轴质流动(axoplasmic flow),其速度为每天1~4mm,主要是将神经元胞体合成的蛋白质,不断地向轴突末端流动,以更新轴质的基质、神经丝以及微管等结构蛋白质。逆向运输(retrograde transport)是轴突末端代谢产物和轴突末端通过人胞作用摄取的蛋白质、神经营养因子以及一些小分子物质等由轴突末端运向胞体,运输方向与轴质流动相反,故称为逆向运输,速度为每天l~4mm,这种运输主要是由多泡体实现。多泡体是一个大泡内含许多小泡,小泡内分别含有代谢产物或摄入的神经营养因子。代谢产物被逆向运输至胞体后,经溶酶体的作用,可分解消化更新,神经营养因子到胞体后,可促进神经元的代谢和调节神经元的生理功能。不论是顺向或逆向运输,均由线粒体提供ATP供能所实现。在某种原因而感染时,有些病毒或毒素由逆向运输,转动到神经元的脑体内而致病。轴突运输是神经元内各种细胞器生理功能的重要体现。轴突的主要功能是将神经冲动由胞体传至其他神经元或效应细胞。轴突传导神经冲动的起始部位,是在轴突的起始段,沿轴膜进行传导。


