神经胶质细胞(神经胶质细胞)

VLoG
次浏览
更新时间:2023-05-23
神经胶质细胞
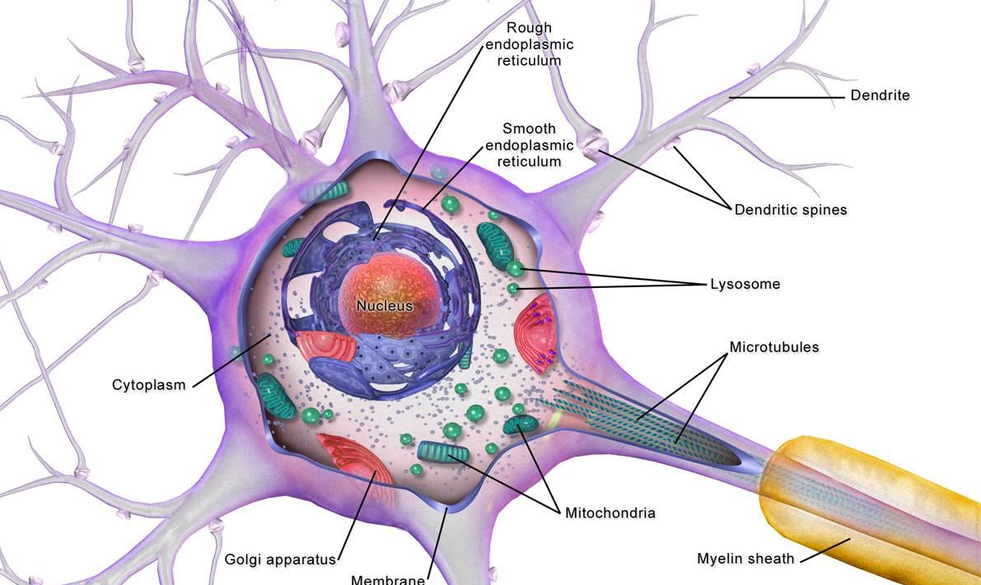
基本信息
| 中文名 | 神经胶质细胞 |
| 外文名 | neuroglial cell |
| 别名 | 胶质细胞 |
| 分布区域 | 中枢和周围神经系统 |
| 种类 | 星形胶质细胞、少突胶质细胞和小胶质细胞 |
| 特征 | 有突起,但无树突和轴突之分 |
| 英文别名 | glial cell |
收起
组织学特点
星形胶质细胞
胶质纤维酸性蛋白(GFAP)分子量为50~52kD,主要存在于中枢神经系统星形胶质细胞。编码GFAP基因位于17q21,与其他中间丝序列尤其是结蛋白和波形蛋白的碱基序列具有较高的同源性,其中与结蛋白的同源性达65%,与波形蛋白的同源性达67%。星形胶质前体细胞主要表达Vimentin,而成熟之后表达Vimentin和GFAP,故GFAP被认为是星形胶质细胞成熟的标志物。有研究发现GFAP阳性细胞在产后小鼠和成年大鼠的中枢神经细胞中呈深染的不规则“星”状,主要定位在星形胶质细胞的胞体和突起上,在前额皮质的分子层、外颗粒层和锥体细胞层、海马和黑质中密集分布,纹状体只有少量阳性细胞表达;在脊髓的灰、白质主要分布于脊髓中央管附近大量表达。在人脑中,GFAP最早出现于胚胎发育的第8周,逐渐表达于胶质细胞中,主要表达于星形胶质细胞中,而在室管膜细胞、少突胶质细胞等胶质细胞也有少量分布。另外,GFAP还可表达在非中枢神经系统(CNS)细胞如许旺细胞、成纤维细胞、肝星状细胞。GFAP蛋白作为成熟的星形胶质细胞中间丝的组成部分,具有调节细胞代谢、形成和维护血脑屏障、产生和释放神经营养因子等作用,而且在维持星形细胞形态和功能上具有重要作用:如形成细胞核和细胞膜的连接、参与细胞骨架重组、黏附和稳定神经元的结构、维持脑内髓鞘形成并作为细胞信号转导通路等。
少突胶质细胞
O1/O4有研究通过间接免疫荧光法观察O1、O2、O3以及O4抗原标记产后早期的小鼠小脑、大脑、脊髓、视神经、视网膜细胞中发现在星形胶质细胞、神经细胞、成纤维细胞的表面都没有检测O抗体,而在少突胶质细胞的表面上检测到,并且O抗原在小鼠、大鼠、鸡和人类的中枢神经系统中均有表达。O4及O1抗原均为少突胶质细胞标志物,均为膜表面的一种脑硫酯,O4既可以表达于晚期少突胶质细胞前体细胞,也可表达于未成熟少突胶质细胞,Q1主要表达于未成熟少突胶质细胞。O4阳性的细胞可以成为分化为成熟MBP阳性的少突胶质细胞,该细胞为GC阳性细胞(少突胶质前体细胞),故可推测O4也特异性表达少突胶质前体细胞。几乎所有的O4阳性的细胞在成熟的大脑皮质也表达NG2(胶质前体细胞),而NG2阳性细胞的不表达OX-42单克隆抗体,说明O4/NG2阳性细胞不是小胶质细胞。为了研究O4是否只在少突胶质细胞谱系表达,进行了O4和GFAP、NFP抗体免疫双标检测,发现O4在皮质灰质中的NFP阳性的神经元细胞、GFAP阳性的星形胶质细胞均不表达。因此,认为O4抗体和GC、NG2一样,可作为少突胶质前体细胞的标志物,但还可标记未成熟少突胶质细胞,不标记神经元细胞、星形胶质细胞、小胶质细胞;而O1仅为未成熟少突胶质细胞的标志物。
少突胶质细胞因子(OLIG)是一个与碱性螺旋-环-螺旋结构转录因子家族,用Northernblot技术分析表明,OLIG1(2.2kb)和OLIG2(2.4kb)在大脑中特异性表达。在成年啮齿动物脑组织,OLIG1/2在少突胶质细胞特异性表达,而在星形胶质细胞和神经元细胞不表达。OLIG1和OLIG2位于鼠16号染色体,在人类位于21号染色体。这两个基因起始于胚胎期脊髓腹侧区域,在胚胎期神经管、脊髓、丘脑均有表达,成年后在胼胝体、脑白质、海马中也可广泛表达,但灰质表达很少。OLIG2的分布要比OLIG1范围广,在脑室区(VZ)、SVZ横向(LGE)区和内侧神经节的E10.5到E14.5区,OLIG2均可呈阳性表达,而在这些细胞中很少表达OLIG1,这表明OLIG2的表达不限于少突胶质细胞系,在多种类型的神经元祖细胞中均可表达,包括多能神经元、神经胶质祖细胞。OLIG1在胚胎期位于少突胶质细胞及其前体细胞的细胞核中,出生后则由胞核迁移到胞质中,但在发生髓鞘损伤后OLIG1又出现在胞核内;而OLIG2始终存在于胞核中,不会发生迁移现象。利用基因转染技术使OLIG1和OLIG2基因过表达,发现过度表达能增加少突胶质前体细胞的增殖和运动神经元前体细胞的产生,其中OLIG1作用于少突胶质细胞的成熟过程中;OLIG2在脊髓和后脑的躯体运动神经元发育过程中发挥作用,也参与少突胶质细胞的发育;OLIG1在发育的脑中能够部分代偿OLIG2的功能。
髓鞘碱性蛋白(MBP)成熟少突胶质细胞的标志物,广泛存在于少突胶质细胞及其髓鞘中的骨架蛋白中,具有四个主要类型,根据分子量分为21500kb,18500kb,17000kb,14000kb。人和鼠的MBP基因均位于第18号染色体远端即18q22-23。MBP分中枢性和周围性两种,中枢性MBP由少突胶质细胞合成和分泌,在白质中含量最高,主要在少突胶质细内以共价键结合于髓鞘质浆膜面,同时也与细胞骨架、微管、微丝相连;周围性MBP由雪旺细胞合成和分泌,存在于周围神经髓鞘中;除了神经组织外,心、肝、肾、肾上腺、骨骼肌等器官组织中也存在,但含量很低,难以测出。在新生小鼠脑的神经胶质细胞培养中采用免疫荧光法和原位杂交的方法研究髓鞘碱性蛋白发育阶段MBP基因的表达,大量MBP特异性mRNA在产后5~6d不成熟少突胶质细胞内突然出现,在成熟过程中慢慢增加,约有60%~80%的细胞可表达MBP,但在神经元细胞中没有表达。有研究用双重标记的方法发现,在小鼠产后8~9d,MBP阳性表达在少量GC阳性的细胞,在产后10~13d,所有的GC阳性细胞均MBP标记阳性,在产后13~14d的细胞培养中MBP标记的细胞比例最高,GC是特异性不成熟的少突胶质细胞的标志物,MBP在GC阳性后的细胞表达,有力地说明MBP表达成熟少突胶质细胞。因此,认为MBP特异性标记少突胶质细胞,而非神经元细胞。
小胶质细胞
CD11b单克隆抗体(OX42)特异性标记小胶质细胞,因能识别鼠的CR3受体,CR3受体位于小胶质细胞的分支,故OX42抗体特异性标记小胶质细胞。在正常成年脑组织中,小胶质细胞处于静息状态,胞体很小,突起很细,OX42染色呈阴性或弱阳性,称之为“静息型”小胶质细胞。当机体受到某种刺激(如外伤、感染、物理化学或电刺激)后小胶质细胞被激活,早期胞体变大、突起变粗、棘明显清晰,OX42染色较深成为早期反应状态,称之为“早期反应型”小胶质细胞。随着刺激灶的存在,小胶质细胞的胞体进一步肥大,突起回缩变短,而成为巨噬细胞样的“吞噬型”小胶质细胞。在年轻组、老年组正常脑组织切片中没有OX42的阳性小胶质细胞,而在大鼠脑出血组中,老年组活化的OX42阳性的小胶质细胞明显增多,而且多于年轻组。故认为OX42单克隆抗体在正常小胶质细胞染色浅或无,而在非正常小胶质细胞染色多或深。
离子钙结合衔接分子1(IBA-1)是一个EF手型蛋白,特异性表达单核细胞谱系,如小胶质细胞。IBA-1是一个具有肌动蛋白属性进化的保守蛋白,已发现与F-肌动蛋白存在共定位,在巨噬细胞集落刺激因子的膜褶皱样运动和吞噬作用中发挥重要作用。IBA-1在人、猴、马、大鼠和小鼠组织的小胶质细胞均有发现。有研究用免疫细胞化学和激光共聚焦显微镜的方法检测IBA-1蛋白只存在小胶质细胞、脑脊膜、室管膜的巨噬细胞和脉络丛浅表的基质细胞的巨噬细胞中表达,而这些细胞均具有吞噬功能,小胶质细胞为巨噬细胞的一种,进一步证实IBA-1在中枢神经系统中特异性表达于小胶质细胞。因IBA-1在小胶质细胞含量丰富且稳定,故认为是小胶质细胞的可靠的特异的标志物。另外,IBA-1也是同种异体移植炎症因子-1(AIF1),还可在造血细胞中表达。
相关视频
全部
457次播放02:00
胶质细胞功能不容小觑,它是中枢神经系统的常驻免疫细胞


