原位杂交(用于基因定位的技术)

VLoG
次浏览
更新时间:2023-05-19
原位杂交
本词条是多义词,共3个义项
用于基因定位的技术

概述
将标记的核酸探针与细胞或组织中的核酸进行杂交,称为原位杂交!使用DNA或者RNA探针来检测与其互补的另一条链在细菌或其他真核细胞中的位置。
RNA原位核酸杂交又称RNA原位杂交组织化学或RNA原位杂交。该技术是指运用cRNA或寡核苷酸等探针检测细胞和组织内RNA表达的一种原位杂交技术。其基本原理是:在细胞或组织结构保持不变的条件下,用标记的已知的RNA核苷酸片段,按核酸杂交中碱基配对原则,与待测细胞或组织中相应的基因片段相结合(杂交),所形成的杂交体(Hybrids)经显色反应后在光学显微镜或电子显微镜下观察其细胞内相应的mRNA、rRNA和tRNA分子。RNA原位杂交技术经不断改进,其应用的领域已远超出DNA原位杂交技术。尤其在基因分析和诊断方面能作定性、定位和定量分析,已成为最有效的分子病理学技术,同时在分析低丰度和罕见的mRNA表达方面已展示了分子生物学的一重要方向。

原位杂交
原理
实验步骤
质粒制备
1质粒的转化和扩增
1.取400uLXL1-Blue菌种加入到含200mlLB培养基的锥形瓶中,37℃、100rpm培养4h,离心,倒置,以冰冷的0.1mol/LCaCl_2重悬细菌,冰浴30min,离心,弃上清,倒置,再加4ml(含15%甘油)冰冷的CaCl2重悬细菌,分装(200μ/tube),-80℃保存。
2.转化:在冰浴中将1管XL1-Blue感受态菌解冻,将浓度为2ng/μ1的质粒DNA4μ1加入到8Oμ1感受态菌中。
3.轻轻摇匀,冰浴30min。
4.42℃热激9O秒,然后迅速冰浴2min。
5.加入LB培养液(无氨苄青霉素)0.8ml,在37℃,100转/min水浴孵育60min。
6.取200μl菌液铺于琼脂板上(涂有X-Gal(20mg/ml)-IPTG(200mg/ml)的LB-氨苄青霉素50mg/ml,1μl/ml培养基),待菌液全部被吸收后,倒置平板于37℃培养12-16h。
3.在70℃温育5min,然后冷却到室温。
4.加1.5μ14mol/LKCl和0.5μ1含0.4%溴酚兰染液,振荡3O秒后,冰浴5min。
5.12000g,4℃离以3min,以除去细菌碎片。
1.3质粒的扩增和纯化
1.用无菌牙签分别挑取单个白色菌落移入含30mlLB-氨苄青霉素(50μg/ml)培养液的聚丙烯管中,于37℃,200转/分培养3h。
2.将菌液转入含70mlLB-氨苄青霉素培养液的250ml锥形瓶中,37℃,200转/分培养过夜(12-16h),细菌浑浊。
4.将培养的细菌倒入50ml的离心管中,6000rpm,4℃离,th,15min,沉淀细菌。
5.弃净上清夜,用2ml预冷的溶液I,悬浮菌体沉淀,剧烈振荡,于室温静置5min
6.加入新配制的溶液II4ml,快速用手晃动10秒,颠倒数次后,于室温静置10min。
7.加入预冷的溶液III3ml,温和振荡l0秒,于冰上静置10min,出现白色絮状沉淀。
8.6000rpm,4℃离心15min,保留上清。
10.12000rpm,4℃离心15min,回收核酸。小心弃去上清,倒置离心管使残兼上清液流尽。
11.于室温用70%的乙醇洗涤沉淀物和管壁,室温12000rpm离心,15min,充分弃去乙醇,于室温将离心管倒置在纸巾上,使最后残余的痕量乙醇挥发殆尽。
12.用500μlTE(pH8.0)溶解核酸沉淀,转移至1.5mlEppendorf管中。
13.加入用冰预冷的5mol/L的LiCl溶液600μl,充分混匀。12000rpm,4℃离心15min,以沉淀高分子量的RNA。
14.将上清转移到另一1.5mlEppendorf管中,加等量异丙醇,充分混匀,于室温静置10min。
15.12000rpm,4℃离心15min,回收核酸。小心弃去上清,倒置离心管使残余上清液流尽。
16.于室温用70%的乙醇洗涤沉淀物和管壁,室温12000rpm离心,15min,充分弃去乙醇,于室温将离心管倒置在纸巾上,使最后残余的痕量乙醇挥发殆尽。
21.于4℃12000rpm离心5min回收沉淀的质粒DNA。尽可能弃去上清,敞开管口,置工作台上使残留的痕量乙醇蒸发殆尽。
22.加入400μl处于4℃的70%乙醇,稍加振荡,漂洗沉淀,4℃12000rpm离心2min。
23.吸去上清,室温敞开管口,直到乙醇完全挥发。
24.用100μlTE缓冲液(pH8.0)溶解沉淀。
25.取4μl溶液1:100稀释后,测定其OD260、OD280,以确定质粒DNA的纯度和浓度(OD260/OD280>1.8。OD260/OD280对DNA而言其值大约为
26.质粒DNA溶液于-20℃保存待用。
探针标记
3.进行体外转录,步骤如下:
质粒DNA模板0.05μg/μl1μl
10xNTP地高辛标记混合物1x2μl
0.1MDTT溶液10mM2μl
RNAse抑制剂2U/μl1μl
RNA聚合酶2U/μl2μl
反应体系总体积20μl
5.加入上述各试剂后,混匀,简短离心后在37℃孵育2h。
6.加入2μl无RNA酶的DNA酶I(10U/μ1),37℃孵育15min降解模板DNA。
8.加入2.5μl的4MLicl和7.5μl冷的无水乙醇,混匀,-20℃放置2h。
9.12000g下离以15min,弃上清,小心地用50μl冷的70%乙醇洗涤沉淀。
10.室温下稍干燥,溶于100μ1DEPC处理过的三蒸水中,混匀分装,-20℃下保持备用。
步骤
3.1冰冻切片与杂交前预处理
1.将子宫样品从-80℃取出,用OCT包埋,在-23℃(切片机腔体温度)平衡至少30min。将包埋好的样品固定在样品头上,切10μm厚的连续组织切片,平铺于涂有多聚赖氨酸(1mg/m1)的玻片上(玻片预先经180℃干烤6小时),保存于-70℃冰柜备用。
2.冰冻切片经室温干燥10min后,在4%的多聚甲醛-PBS(pH7.4)固定l0min。
5.在切片上滴加蛋白酶K(0.1μg/m1),37℃孵育15min,重复步骤4)。
7.5xSSC中平衡15min。
3.2杂交
8.在脱水后的玻片上滴加预杂交液(约100μl/玻片),置于放有湿盒液(50%甲酰胺v/v;0.3MNaCl;1MmEDTA;10mMTris-Cl,pH8.O)的湿盒中,55~58℃下的烘箱中预杂交2h。
9.甩掉预杂交液,地高辛标记的反义或正义cRNA探针(浓度1-2ng/μl)经70℃变性1O分钟,置冰上1min,玻片上滴加预杂交液(约60μl/玻片),覆盖parafilm膜,放湿盒中在48~58℃下杂交18-30h。
3.3杂交后处理
10.取出玻片,小心去掉Parafilm膜,甩掉杂交液,用52℃预热的5×SSC洗30min。
11.在无DNA的RNA酶A(20μg/ml)溶液中37℃下孵育30min。
12.分别依次用52℃预热的2XSSC,1XSSC和O.1XSSC洗2次,每次30min。
3.4杂交信号检测
13.在缓冲液A(0.1MTris-HC1pH7.5,0.15MNaC1)中平衡5min。
14.在玻片上滴加碱性磷酸酶的抗地高辛抗体(1:500~1:2000稀释于含0.5%阻断液的缓冲液A中),室温反应2h。
15.用缓冲液A洗2次,每次15min。
18.充分显色后,用EDTA(1mMEDTA,pH8.0)洗15min终止反应。
19.在95%乙醇中洗lh以除去非特异的背景。
20.用蒸馏水洗15min除去可能存在的结晶体。
22.充分干燥后在显微镜下观察、照相。
前固定
原位杂交组织化学技术(InSituHybridizationHistochemistry,ISHH)在固定剂的应用和选择上应兼顾到三个方面:保持细胞结构,最大限度地保持细胞内DNA或RNA的水平;使探针易于进入细胞或组织。DNA是比较稳定的,mRNA是相对稳定的但易为酶合成和降解。RNA却绝然不同,非常容易被降解。因此,对于DNA的定位来说,固定剂的种类和浓度并不十分重要。相反,在RNA的定位上,如果要使RNA的降解减少到最低限度,那么,不仅固定剂的种类浓度和固定的时间十分重要,而且取材后应尽快予以冷冻或固定。在解释ISHH的结果时应考虑到取材至进入固定剂或冰冻这段时间对RNA保存所带来的影响,因组织中mRNA的降解是很快的。在固定剂中,最常用的是多聚甲醛。和其它的固定剂(如戊二醛)不同,多聚甲醛不会与蛋白质产生广泛的交叉连接,因而不会影响探针穿透入细胞或组织。
其它如醋酸-酒精的混合液和Bouin’s固定剂也能获得较满意的效果。对于mRNA的定位,我们常采用的方法是将组织固定于4%多聚甲醛磷酸缓冲液中1~2h,在冷冻前浸入15%蔗糖溶液中,置4℃冰箱过夜,次日切片或保存在液氮中待恒冷箱切片机或振荡切片机切片。组织也可在取材后直接置入液氮冷冻,切片后才将其浸入4%多聚甲醛约10min,空气干燥后保存在-70℃。如冰箱温度恒定,在-70℃可保存数月之久不会影响杂交结果。在病理学活检取材多用福尔马林固定和石蜡包埋,这种标本对检测DNA和mRNA有时也可获得杂交信号,但石蜡包埋切片由于与蛋白质交叉连接的增加,影响核酸探针的穿透,因而杂交信号常低于冰冻切片。同时,在包埋的过程中可减低mRNA的含量。
其它固定剂如应用多聚甲醛蒸汽固定干燥后的冷冻切片也可获得满意效果。各种固定剂均有各自优缺点,如沉淀性(Precipitating)固定剂:酒精/醋酸混合液、Bouin’s液、Carnoy’s液等能为增加核酸探针的穿透性提供最佳条件,但它们不能最大限度地保存RNA,而且对组织结构有损伤。戊二醛能较好地保存RNA和组织形态结构,但由于和蛋白质产生广泛的交叉连接,从而大大地影响了核酸探针的穿透性。至今,多聚甲醛仍被公认为ISHH较为理想的固定剂。
处理
1.玻片的处理。玻片包括盖片和载片应用热肥皂刷洗,自来水清洗干净后,置于清洁液中浸泡24h,清水洗净烘干,95%酒精中浸泡24h后蒸馏水冲洗、烘干,烘箱温度最好在150℃或以上过夜以去除任何RNA酶。盖玻片在有条件时最好用硅化处理,锡箔纸包裹无尘存放。由于ISHH的实验周期长,实验程序繁杂,因此,要应用粘附剂预先涂抹在玻片上,干燥后待切片时应用,以保证在整个实验过程中切片不致脱落。常用的粘附剂有铬矾-明胶液,其优点是价廉易得,但在长周期实验过程中,粘附效果不够理想。多聚赖氨酸液具有较好的粘附效果,但价格昂贵,需进口。
2.增强组织的通透性和核酸探针的穿透性。此步骤根据应用固定剂的种类、组织的种类、切片的厚度和核酸探针的长度而定。比如用戊二醛固定的组织由于其与蛋白质产生广泛的交叉连接就需要应用较强的增强组织通透性的试剂。增强组织通透性常用的方法如应用稀释的酸洗涤、去垢剂(detergent)或称清洗剂TritonX-100、酒精或某些消化酶如胃蛋白酶、胰蛋白酶、胶原蛋白酶和淀粉酶(diastase)等。这种广泛的去蛋白作用无疑可增强组织的通透性和核酸探针的穿透性,提高杂交信号,但同时也会减低RNA的保存最和影响组织结构的形态,因此,在用量及孵育时间上应慎为掌握。
蛋白酶K(ProteinaseK)的消化作用在ISHH中是应用于蛋白消化的关键步骤,其浓度及孵育时间视组织种类、应用固定剂种类、切片的厚薄而定。一般应用酶K1μg/ml(于0.1mol/LTris/50mmol/LEDTA,pH8.0缓冲液中),37℃孵育15~20min,以达到充分的蛋白消化作用而不致影响组织的形态为目的。蛋白酶K还具有消化包围着靶DNA的蛋白质的作用,从而提高杂交信号。在蛋白酶K消化后,应用0.1mol/L的甘氨酸溶液(在PBS中)清洗以终止蛋白酶K的消化作用,甘氨酸是蛋白酶K的抑制剂。为保持组织结构,通常用4%多聚甲醛再固定。Burns等(1987)报告应用胃蛋白酶(pepsin)20~100μg/ml(用0.1NHCl配)37℃、30min进行消化,所获实验结果优于蛋白酶K。
不少实验工作者在多聚甲醛固定后,浸入乙酸酐(aceticanhydride)和三乙醇胺(tri-ethanolamine)中以减低静电效应,减少探针对组织的非特异性背景染色。有的作者除在室温下浸于上述溶液10min外,还在预热37℃的50%甲酰胺/2×SSC液中预杂交15min,然后用2×SSC,0.30mol/LNaAc/0.030mol/L枸橼酸钠液中浸15min。但Heinz、Hofer等一些著名学者却对此持有异议,根据他们的实验和经验证明,乙酸酐和三乙醇胺液的处理并不能起到减低背景的目的,不能改善ISHH的信/噪比例。
3.减低背景染色。和免疫细胞化学染色一样ISHH实验程序中,如何减低背景染色是一个重要的问题。ISHH中背景染色的形成是诸多因素构成的。杂交后(Posthybridization)的酶处理和杂交后的洗涤均有助于减低背景染色。预杂交(Prehybridization)是减低背景染色的一种有效手段。预杂交液和杂交液的区别在于前者不含探针和硫酸葡聚糖(Dextransulphate)。将组织切片浸入预杂交液中可达到封闭非特异性杂交点的目的,从而减低背景染色。有的实验室在杂交后洗涤中采用低浓度的RNA酶溶液(20μg/ml)洗涤一次,以减低残留的和内源性的RNA酶,减低背景染色。
4.防止RNA酶的污染。由于在手指皮肤及实验用玻璃器皿上均可能含有RNA酶,为防止其污染影响实验结果,在整个杂交前处理过程都需戴消毒手套。所有实验用玻璃器皿及镊子都应于实验前一日置高温(240℃)烘烤以达到消除RNA酶的目的。要破坏RNA酶,其最低温度必须在150℃左右。有条件的国外实验室在消毒的玻璃器皿外包以锡箔纸以利于标记和防止取出时空气污染。在无高温消毒的烤箱时,亦可用国内出产的卫生蒸汽消毒锅(山东新华医疗器材厂生产)。杂交前及杂交时所应用的溶液均需经高压消毒处理。
过程
杂交(Hybridisation),在ISHH,整个实验周期是比较长的,实验程序也比较繁杂,而杂交在ISHH整个实验中可被认为是“短兵相接”的一步。杂交前的一切准备工作如增加组织通透性都是为了在杂交这一步骤中核酸探针能进入细胞或组织与其内的靶核苷酸相结合。因此,杂交是ISHH中关键的而且是最重要的一个环节。
杂交是将杂交液滴于切片组织上,加盖硅化的盖玻片。国内向正华等采用无菌的蜡膜代替硅化的盖玻片也可获得满意的实验结果。加盖片的目的是防止孵育过程中的高温(50℃左右)导致杂交液的蒸发。因此,也有为稳妥起见,在盖玻片周围加液体石蜡封固的,但作者认为这并不十分必要,因封固的石蜡在高温下融解反易导致杂交液的污染,必要时可加橡皮泥封固盖片四周。硅化的盖玻片的优点是清洁无杂质,光滑不会产生气泡和影响组织切片与杂交液的接触,盖玻片自身有一定重量能与有限的杂交液吸附达到覆盖和防止蒸发的作用。在孵育时间较长时,为保证杂交所需的湿润环境,可将复有硅化盖玻片进行杂交的载片放在盛有少量5×SSC或2×SSC(standardsalinecitrate,SSC)溶液的硬塑料盒(要能防止高温破坏)中进行孵育。杂交液的成分和预杂交液基本相同,所不同的是加入了标记的核酸探针和硫酸葡聚糖。
如前所述,杂交前的准备只是为杂交的成功奠定基础,要获得满意的实验结果,在杂交这一实验过程中还须注意以下的环节。
①探针的浓度。很难事先确定每一种实验探针的浓度,但要掌握一个原则,即探针浓度必须给予该实验最大的信/噪比值。背景染色的高低也与探针浓度有关。根据国内外实验工作者的经验,认为最佳原则应是应用最低探针浓度以达到与靶核苷酸的最大饱和结合度为目的。这和我们在免疫细胞化学试验中选择抗血清的最佳工作浓度的原则一样。探针浓度依其种类和实验需要略有不同,根据笔者的经验及所查阅文献,在原位杂交细胞化学中,探针浓度为0.5~5.0μg/ml(即0.5~5.0ng/μl)。
根据Heinz、Hofelt实验室经验,对放射性标记的dsDNA或cRNA探针,其浓度在2~5ng/μl。Conlton认为生物素标记探针,其最佳浓度在0.5~5ng/μl。作者在英皇家研究生院Polak教授实验室应用于放射性标记cRNA探针的浓度为0.5ng/μl,而在非放射性标记(生物素或地高辛)cRNA探针浓度为2.5ng/μl,放射性标记DNA探针浓度为1.0ng/μl。向正华等应用地高辛标记生长抑素cRNA探针获得满意结果,其探针浓度为0.5ng/μl。必须强调的是,国内外实验室都证明加杂交液的量要适当,以10~20μl/每张切片为宜。杂交液过多不仅造成浪费,而且液量过多常易致盖玻片滑动脱落,影响杂交效果,过量的杂交液含核酸探针浓度过高,反易导致高背景染色等不良后果。
②探针的长度。一般应用于ISHH探针的最佳长度应在50~100个碱基之间。探针短易进入细胞,杂交率高,杂交时间短。据报告,长500个碱基的探针,其杂交时间约需20h左右。200~500个碱基的探针仍可应用,如超过500个碱基的探针则在杂交前最好用碱或水解酶进行水解,使其变成短的片段,达到实验所需求的碱基数。
③杂交的温度和时间。杂交的温度也是杂交成功与否的一个重要环节。在第十八章概述中曾提到DNA或RNA需加热或变性、解链后才能进行杂交。能使50%的核苷酸变性解链所需的温度,叫解链温度或融解温度(meltingtemperature,简称Tm)。原位杂交中,多数DNA探针需要的Tm是90℃,而RNA则需要95℃。这种高温对保存组织形态完整和保持组织切片粘附在载玻片上是不可能的。因此,在杂交的程序中常规的加入30%~50%甲酰胺(for-mamide)于杂交液中。McConaughy报告,反应液中每增加1%的甲酰胺浓度,Tm值可降低0.72℃。因此,可用调节盐浓度的办法来调节Tm。Tm的计算公式在第十九章有介绍,由公式的列出也表明了它与盐的浓度、探针的长度、甲酰胺的百分比等诸多因素有关。
由于盐和甲酰胺浓度的调节等因素,实际采用的原位杂交的温度在Tm-25℃左右,即比Tm减低25℃,大约在30~60℃之间,根据探针的种类不同,温度略有差异,RNA和cRNA探针一般在37~42℃左右,而DNA探针或细胞内靶核苷酸为DNA的,则必须在80~95℃加热使其变性,时间5~15min,(有作用报告在105℃微波炉加热使之变性),然后在冰上搁置1min,使之迅速冷却,以防复性,再置入盛有2×SSC的温盒内,在37~42℃孵育杂交过夜。杂交的时间如过短会造成杂交不完全,而过长则会增加非特异性染色。从理论上讲,核苷酸杂交的有效反应时间在3h左右。Barns等(1987)报告用DNA探针杂交,其反应完成时间为2~4h。但为稳妥起见,一般将杂交反应时间定为16~20h,或为简便起见杂交孵育过夜,从现有文献报告看无不良结果。当然,杂交反应的时间与核酸探针长度与组织通透性有关,在确定杂交反应时间应予考虑,并经反复实验确定。有作者主张杂交反应的孵育应在黑暗环境中进行,因为光线会促进甲酰胺的电离作用。
④杂交严格度(Hybridizationstringency)。杂交条件的严格度(stringency)表示通过杂交及冲洗条件的选择对完全配对及不完全配对杂交体的鉴别程度。错配对(mismatch)杂交的稳定性较完全配对杂交体差,因此,通过控制杂交温度、盐浓度等,可减弱非特异性杂交体的形成,提高杂交的特异性。所以,杂交的条件愈高,特异性愈强,但敏感性降低,反应亦然。一般来说,低严格度(lowstringency)杂交及冲洗条件在Tm-35℃至Tm–40℃之间,高盐或低甲酰胺浓度。在这种条件下,大约有70%~90%的同源性核苷酸序列被结合,其结果是导致非特异性杂交信号的产生。中严格度,Tm-20℃至Tm-30℃的范围。高严格度(highstringency)为Tm-10℃至Tm-15℃,低盐和高甲酰胺浓度。
在这种条件下,只有具有高同源性的核苷酸序列才能形成稳定的结合。麦跃行装用地高辛标记原位杂交技术检测尖锐湿疣中人乳头瘤病毒DNA型别,结果发现在严格条件下(Tm-12℃)各型病毒DNA的检出率和阳性率明显低于非严格条件下(Tm-35℃),其相差非常明显(P<0.001)。因为,在严格条件下只有同源性很强的DNA才被检出,而在非严格条件下同源性较低的DNA序列也被检出。因此,他建议对病毒DNA分型需在高严格条件下进行,而低严格条件则可用于对病毒感染进行筛选。由于原位杂交技术多数是在Tm-25℃进行的,不属于高严格范围,无疑会产生非特异性结合导致信/噪比减低。在这种情况下,可用加强杂交后处理洗涤的严格度使非特异性的杂交体减少。
由于RNA杂交的稳定性,应用cRNA探针进行细胞或组织的原位杂交时的杂交温度比其它核酸探针要高10~15℃。实验证明,cRNA产生的信号比双链cDNA要强。单链的RNA探针其杂交信号大于双链的cDNA的约8倍。
⑤硫酸葡聚糖(Dextransulphate)和甲酰胺(formamide)。硫酸葡聚糖是核酸杂交液中仅次于甲酰胺的一种组成成份。在杂交液中,甲酰胺占50%左右,而硫酸葡聚糖占10%左右。它是一种大分子的多聚胺化合物,具有极强的水合(hydrate)作用,因而能大大增加杂交液的粘稠度。硫酸葡聚糖的主要作用是促进杂交率,特别是对双链核酸探针。这是应用硫酸葡聚糖于杂交液中的主要目的。甲酰胺的主要作用在上节已提及,在调节杂交反应温度方面,甲酰胺起了极为重要的作用,从而有助于保持组织的形态结构。甲酰胺还可防止在低温时非同源性片段的结合,但甲酰胺具有破坏氢键的作用从而具有一种不稳定的作用。
后处理
杂交后处理包括系列不同浓度,不同温度的盐溶液的漂洗。在原位杂交组织化学的实验程序中,这也是一个重要的环节。特别因为大多数的原位杂交实验是在低严格度条件下进行的,非特异性的探针片段粘附在组织切片上,从而增强了背景染色。RNA探针杂交时产生的背景染色特别高,但能通过杂交后的洗涤有效地减低背景染色,获得较好的反差效果。在杂交后漂洗中的RNA酶液洗涤能将组织切片中非碱基配对RNA除去。洗涤的条件如盐溶液的浓度、温度、洗涤次数和时间因核酸探针的类型和标记的种类不同而略有差异,一般遵循的共同原则是盐溶液浓度由高到低而温度由低到高。必须注意的是在漂洗的过程中,切勿使切片干燥。干燥的切片即使大量的溶液漂洗也很难减少非特异性结合,从而增强了背景染色。放射性标记探针杂交后漂洗过程中可用底片曝光的方法检测背景染色(非特异性标记的多少)作为改善漂洗程序的指针。在35S标记的核酸探针在漂洗液中须加入14mmol/L的β-巯基乙醇(β-mercaptoethanol)或硫代硫酸盐(thiosulphate),以防止35S标记的核酸探针被氧化。总之,如何控制漂洗的严格度从而达到理想的信/噪比无既定方案可循,必须从反复的实践中取得经验。
显示
显示又可称为检测系统(Detectionsystem)。根据核酸探针标记物的种类分别进行放射自显影或利用酶检测系统进行不同显色处理。细胞或组织的原位杂交切片在显示后均可进行半定量的测定,如放射自显影可利用人工或计算机辅助的图象分析检测仪(computer–assistedimageanalysis)检测银粒的数量和分布的差异。非放射性核酸探针杂交的细胞或组织可利用酶检测系统显色,然后利用显微分光光度计或图像分析仪对不同类型和数量的核酸的显色强度进行检测。但利用ISHH做半定量测定必须注意严格控制实验的同一条件,切片的厚度和核酸的保存量如取材固定的间隔时间等。如为放射自显影,核乳胶膜的厚度与稀释度等必须保持一致。
例子
对分散在若干个琼脂平板上的少数菌落(100-200)进行克隆筛选时,可采用该方法。将这些菌落归并到一个琼脂主平板以及已置于第二个琼脂平板表面的一张硝酸纤维素滤膜上。经培养一段时间后,对菌落进行原位裂解。主平板应贮存于4℃直至得到筛选结果。
1.将少数菌落转移到硝酸纤维素滤膜上
(2)用无菌牙签将各个菌落先转移至滤膜上,再转移至含有选择性抗生素但未放滤膜的琼脂主平板上。应按一定的格子进行划线接种(或打点)。每菌落应分别划线于两个平板的相同位置上。最后,在滤膜和主平板上同时划一个含有非重组质粒(如pBR322)的菌落。
(5)用Parafilm膜封好主平板,倒置贮放于4℃,直至获得杂交反应的结果。
(6)裂解细菌,按本段下面所述方法,使释放的DNA结合于硝酸纤维素滤膜。
2.菌落的裂解及DNA结合于硝酸纤维素滤膜
(1)在一张保鲜膜上制作一个装有0.5mol/LNaOH的小洼(0.75ml),使菌落面朝上,将滤膜放到小洼上,展平保鲜膜,使滤膜均匀湿润,让滤膜留于原处2-3分钟。 (2)用干纸巾从滤膜的下方吸干滤膜,用一张新的保鲜膜和新配制的0.5mol/LNaOH重复步骤(1)。
(3)吸干滤膜,将滤膜转移到新的带有1mol/LTris·Cl(pH7.4)的保鲜膜洼上。5分钟后吸干滤膜,再重复一次该步骤。
(4)吸干滤膜,把它转移到有1.5mol/LNaCl、0.5mol/LTris·Cl(pH7.4)的保鲜膜小洼上5分钟后吸干滤膜,转移到一张干的滤纸上,置于室温20-30分钟,使滤膜干燥。 (5)将滤膜夹在两张干的滤纸之间,在真空烤箱中于80℃干烤2小时,固定DNA。
(6)将固定在膜上的DNA与32P标记的RNA进行杂交。
3.杂交
(2)将滤膜转到200ml预洗液的玻璃皿中。滤膜何叠在一起,放于溶液中。用保鲜膜盖住玻璃皿,放到位于培养箱内的旋转平台上。于50℃处理30分钟。在这一步及以后的所有步骤中,应缓缓摇动滤膜,防止它们粘在一起。
(3)用泡过预洗液的吸水棉纸轻轻地从膜表面拭去细菌碎片,以降低杂交背景而不影响阳性杂交信号的强度和清晰度。
(4)将滤膜转到盛有150ml预杂交液的玻璃中,在适宜温度(即在水溶液中杂交时用68℃,而在50%甲酰胺中杂交时用42℃)下,预杂交1-2小时。
(7)68℃用300-500ml1×SSC和0.1%SDS溶液洗膜两次,每次1-1.5小时。此时已可进行放射自显影。如背景很高或实验要求严格的洗膜条件,可用300-500ml0.2×SSC和0.1%SDS的溶液于68℃将滤膜浸泡60分钟。
(8)把滤膜放在纸巾上于室温晾干后,把滤膜(编号面朝上)放在一张保鲜膜上,并在保鲜膜上作几个不对称的标记,以使滤膜与放射性自显影片位置对应。
(9)用第二张保鲜膜盖住滤膜。加X光片并加上增感屏于-70℃曝光12-16小时。
试验
收集斑马鱼的胚胎,在Holfretor水中培养,到达所需要的发育时期时,用蛋白酶去除卵膜,用4%多聚甲醛固定,在4℃保存,二十四小时后用50%甲醇2%多聚甲醛溶液洗,然后换成甲醇,在-20C保存,待用(两天和两天以上的胚胎需要用双氧水处理,去除色素。或者使用苯锍脲稀溶液培养,可阻断色素的形成)
一.原位杂交第一天
1.重新水化和固定
1)吸取固定好的胚胎,加入50%甲醇的PBST溶液,放置5分钟。
2)置换成30%甲醇的PBST溶液,放置5分钟
3)置换成PBST溶液,放置5分钟,重复一次
4)置换成4%多聚甲醛的PBS溶液固定20分钟
5)用PBST洗两次,每次放置五分钟,室温。
蛋白酶处理与后固定(本实验不做此步)
1)用10ul/ml的蛋白酶K在室温下处理胚胎。5体节以下的胚胎不处理,5体节到24小时的胚胎处理3分钟,24小时以上的胚胎处理5分钟或者更长。发育时间越短的胚胎越嫩,可以不用或者少用蛋白酶处理,发育时间长的胚胎就需要用蛋白酶来疏松组织,以便于杂交。
2)用PBST溶液轻洗,在PBST中放置5分钟。
3)用4%多聚甲醛的PBS溶液固定20分钟,室温。
4)用PBST洗两次,每次放置五分钟,室温。
2.预杂交
1)每个管中置换成大约300ulHYB-溶液,60℃水浴5分钟,避免振荡。
2)用等体积的HYB+取代HYB-。
3)60℃水浴,预杂交4小时以上。
3.杂交
1)吸去预杂交的HYB+,加上100ul已加入探针的HYB+溶液(探针浓度约为1ng/ul)..
2)60℃温浴过夜。
注:杂交与预杂交的温度可以是55到60度不等,温度越低,探针结合越好,温度越高,背景越小。
二.原位杂交第二天
1.
1)将探针回收,放于-20C保存(通常探针可重复使用十次左右)。
2)加入50%甲酰胺/2XSSCT溶液1毫升,60℃,放置30分钟,重复一次。
3)置换2XSSCT1ml,60℃,放置15分钟。
4)置换0.2XSSCT1ml,60℃,放置30分钟,重复一次。
2.
1)用MABT洗两次,每次五分钟,放在摇床轻轻摇动。
2)室温下加1ml1:2:7溶液,时间为一小时。
3)按1:3000的比例在1:2:7溶液中加入酶连地高辛抗体,4C冰箱过夜。
三.原位杂交第三天
2)用1ml1mM左旋米睉的Stainingbuffer洗三次,每次放置五分钟。
3)将胚胎转入十六孔板中,吸去stainingbuffer,加上300ulBMPurpleAPSubstrate(底物,用之前加5mM左旋米唑),十六孔板外面包上锡箔纸以避光,避免摇动,室温下显色。
4)每隔一小时观察胚胎是否开始显色
5)将显色完全的胚胎中的底物吸出,用PBST洗两三次后加上4%多聚甲醛固定,拍照。
6)4C冰箱保存。
原位杂交中溶液的配制:
PBS:
NaCl8g
KCl0.2g
Na2HPO41.44g
KH2PO40.24g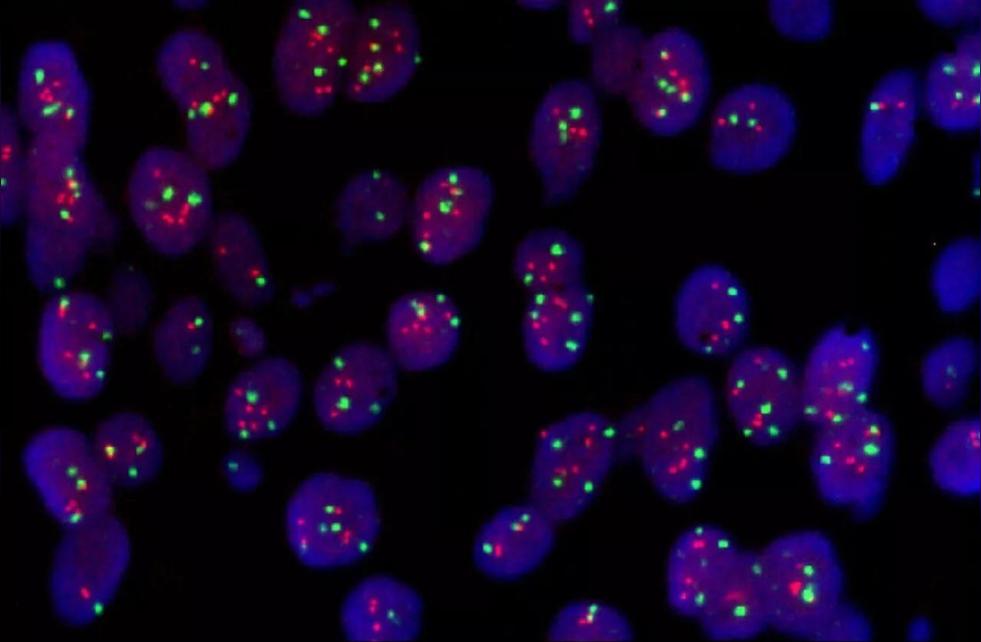

原位杂交
DEPCH2O1L
HCl调PH值至7.4
抽滤,灭菌
4%多聚甲醛:
多聚甲醛40g
PBS1L
加热持续搅拌至溶液澄清。-20℃保存
PBST:
20XSSC:
Na3Citrate2H2O88.2g
NaCl175.5g
DEPCH2O至1L
抽滤,灭菌
SSCT:
SSC加上Tween-20使其终浓度为0.1%。
HYB-:
加入Tween-20使其终浓度为0.1%,-20c保存。
HYB+:
HYB-20ml
yeastRNA10mg
heparin1mg。
-20℃保存。
MAB:
maleicacid11.6g
NaCl8.8g
用固体NaOH(约7g)调至Ph=7.5
4℃保存
MABT
MAB加上Tween-20使其终浓度为0.1%.
10%blockingreagent:
blockingreagent8g
MAB72ml
1:2:7溶液:
灭活羊血清:10%BMblockingreagent:MABT=1:2:7
用时现配
Stainingbuffer:
Tris12.1g
pH9.5
Mgcl6H2O10.2g,
Nacl5.85g
Tween-201ml,
用之前加1M的左旋咪唑储液,使之终浓度为1mM。
应用
意义
原位杂交:在研究DNA分子复制原理的基础上发展起来的一种技术。其基本原理是两条核苷酸单链片段,在适宜的条件下,能过氢键结合,形成DNA-DNA、DNA-RNA或RNA-RNA双键分子的特点,应用带有标记的(有放射性同位素,如3H、35S、32P、荧光素生物素、地高辛等非放射性物质)DNA或RNA片段作为核酸探针,与组织切片或细胞内待测核酸(RNA或DNA)片段进行杂交,然后可用放射自显影等方法予以显示,在光镜或电镜下观察目的mRNA或DNA的存在并定位;用原位杂交技术,可在原位研究细胞合成某种多肽或蛋白质的基因表达。此方法有很高的敏感性和特异性,可进一步从分子水平来探讨细胞的功能表达及其调节机制。已成为当今细胞生物学、分子生物学研究的重要手段。
判断
对照试验和ISHH结果判断,和其它实验方法一样,并非ISHH的任何阳性信号都是特异性的,故必须同时有对照试验以证明其特异性。对照试验的设置须根据核酸探针和靶核苷酸的种类和现有的可能条件去选定。从理论上讲,对照试验设置愈多其靶核苷酸特异性确定愈可靠,但现实是不可能的。因此,在上述对照试验中应任选设至少3~4种用以证实ISHH结果的可靠性。在上述试验中,标明*者为比较可靠的对照试验。
②如果具备相当的免疫组化抗血清,可用结合的免疫组织化学和ISHH法从蛋白质(或多肽)水平和转录水平在相邻切片或同一切片中证明同一种多肽和相应mRNA共存于同一细胞中。
③预先将切片用DNA酶或RNA酶消化,然后用ISHH技术证明丢失的是DNA或RNA。如同免疫组化的吸收试验一样,事先与特异性的cRNA或cDNA进行杂交。再进行ISHH,其结果应为阴性。由于同义RNA探针和组织内mRNA序列顺序是相同的,应用其进行ISHH,结果应为阴性。
④检测系统的对照如乳胶或酶显色系统也应在无标记探针的情况下进行。ISHH的最大优点是它的高度特异性,它可测定组织、培养的单个细胞或细胞提取物中的核苷酸含量。应用高敏感度的放射性标记cRNA探针在理想的ISHH的实验条件下检测mR-NA,其敏感度可达到20个mRNA拷贝/每个细胞。由于双链DNA的稳定性,在用ISHH定位DNA时很少发生丢失,降解。在靶核苷酸序列比较伸展的情况如染色体铺片,长于2kb的探针可以应用。因此,其敏感性高到能够出在染色体铺片上,有时甚至在组织切片上的单个基因拷贝。正因为如此,对ISHH结果的解释应持慎重态度,特别是前人未报告过的新发现。因为如前所述,影响ISHH实验结果的因素太多,比如在外科或实验取材后未及时的固定或冷冻可由于组织中mRNA的降解而导致假阴性结果。另外,在各种类型核酸探针进入细胞、组织和各种器官的能力,又叫可接近性(acessiblity)各异。这些诸多因素都将影响ISHH的实验结果。
荧光
荧光原位杂交(fluorescenceinsituhybridization,FISH)是在20世纪80年代末在放射性原位杂交技术的基础上发展起来的一种非放射性分子细胞遗传技术,以荧光标记取代同位素标记而形成的一种新的原位杂交方法,探针首先与某种介导分子(reportermolecule)结合,杂交后再通过免疫细胞化学过程连接上荧光染料,FISH的基本原理是将DNA(或RNA)探针用特殊的核苷酸分子标记,然后将探针直接杂交到染色体或DNA纤维切片上,再用与荧光素分子偶联的单克隆抗体与探针分子特异性结合来检测DNA序列在染色体或DNA纤维切片上的定性、定位、相对定量分析.FISH具有安全、快速、灵敏度高、探针能长期保存、能同时显示多种颜色等优点,不但能显示中期分裂相,还能显示于间期核。同时在荧光原位杂交基础上又发展了多彩色荧光原位杂交技术和染色质纤维荧光原位杂交技术.
参考资料
[1]
原位杂交染色[引用日期2014-09-15]
[2]
研域(上海)化学试剂有限公司[引用日期2012-01-16]
[3]
价值中国网[引用日期2012-01-16]


