遗传学(研究生物遗传与变异的科学)

VLoG
次浏览
更新时间:2023-05-22
遗传学
本词条是多义词,共6个义项
研究生物遗传与变异的科学
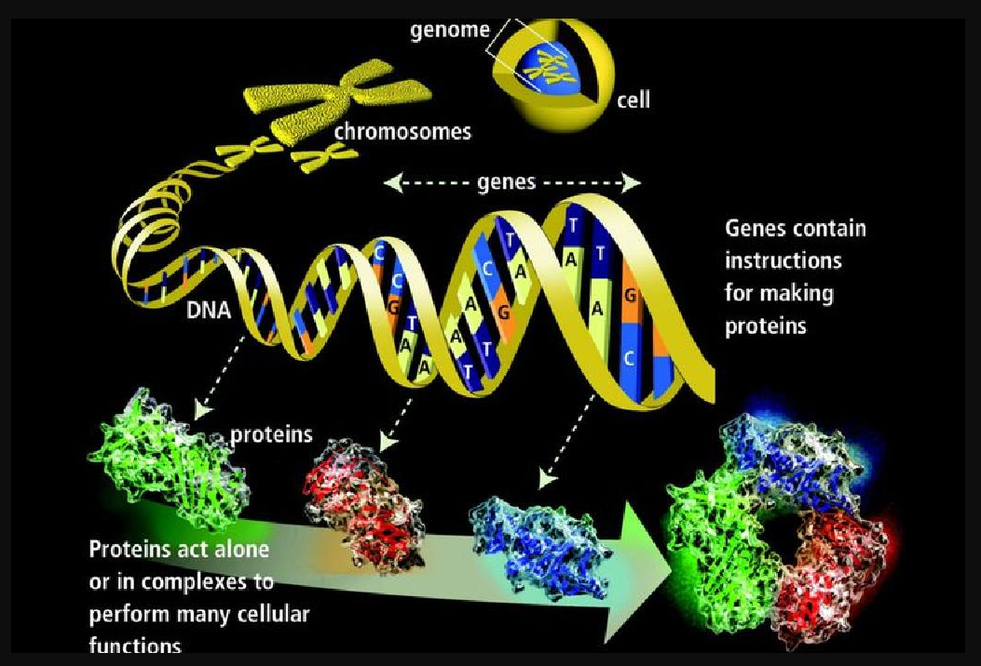
遗传学(Genetics)——自然科学领域中探究生物遗传和变异规律的科学。
研究基因的结构、功能及其变异、传递和表达规律的学科(全国科学技术名词审定委员会审定·遗传学名词。北京:科学出版社,2006年3月)。
基本信息
| 中文名 | 遗传学 |
| 外文名 | Genetics |
| 研究对象 | 父母子女或一个家族 |
| 研究范围 | 遗传与变异的科学 |
| 重要物质 | DNA |
| 单位 | 细胞为单位 |
| 学科代码 | 18031 |
收起
基本要领
学科分支

从噬菌体到人,生物界有基本一致的遗传和变异规律,所以遗传学原则上不以研究的生物对象划分学科分支。人类遗传学的划分是因为研究人的遗传学与人类的幸福密切相关,而系谱分析和双生儿法等又几乎只限于人类的遗传学研究。
更多的遗传学分支学科是按照所研究的问题来划分的。例如,细胞遗传学是细胞学和遗传学的结合;发生遗传学所研究的是个体发育的遗传控制;行为遗传学研究的是行为的遗传基础;免疫遗传学研究的是免疫机制的遗传基础;辐射遗传学专门研究辐射的遗传学效应;药物遗传学则专门研究人对药物反应的遗传规律和物质基础,等等。
从群体角度进行遗传学研究的学科有群体遗传学、生态遗传学、数量遗传学、进化遗传学等。这些学科之间关系紧密,界线较难划分。群体遗传学常用数学方法研究群体中的基因的动态,研究基因突变、自然选择、群体大小、交配体制、迁移和漂变等因素对群体中的基因频率和基因平衡的影响;生态遗传学研究的是生物与生物,以及生物与环境相互适应或影响的遗传学基础,常把野外工作和实验室工作结合起来研究多态现象、拟态等,借以验证群体遗传学研究中得来的结论;进化遗传学的研究内容包括生命起源、遗传物质、遗传密码和遗传机构的演变以及物种形成的遗传基础等。物种形成的研究也和群体遗传学、生态遗传学有密切的关系。
从应用角度看,医学遗传学是人类遗传学的分支学科,它研究遗传性疾病的遗传规律和本质;临床遗传学则研究遗传病的诊断和预防;优生学则是遗传学原理在改良人类遗传素质中的应用。生统遗传学或数量遗传学的主要研究对象是数量性状,而农作物和家畜的经济性状多半是数量性状,因此它们是动植物育种的理论基础。
研究方法
发展简史
成立基础

孟德尔
许多人在这些活动的基础上力图阐明亲代和杂交子代的性状之间的遗传规律都未获成功。直到1866年奥地利学者孟德尔根据他的豌豆杂交实验结果发表了《植物杂交试验》的论文,揭示了现在称为孟德尔定律的遗传规律,才奠定了遗传学的基础。
孟德尔的工作结果直到20世纪初才受到重视。19世纪末叶在生物学中,关于细胞分裂、染色体行为和受精过程等方面的研究和对于遗传物质的认识,这两个方面的成就促进了遗传学的发展。
从1875~1884的几年中德国解剖学家和细胞学家弗莱明在动物中,德国植物学家和细胞学家施特拉斯布格在植物中分别发现了有丝分裂、减数分裂、染色体的纵向分裂以及分裂后的趋向两极的行为;比利时动物学家贝内登还观察到马副蛔虫的每一个身体细胞中含有等数的染色体;德国动物学家赫特维希在动物中,施特拉斯布格在植物中分别发现受精现象;这些发现都为遗传的染色体学说奠定了基础。美国动物学家和细胞学家威尔逊在 1896年发表的《发育和遗传中的细胞》一书总结了这一时期的发现。
关于遗传的物质基础历来有所臆测。例如1864年英国哲学家斯宾塞称之为活粒;1868年英国生物学家达尔文称之为微芽; 1884年瑞士植物学家内格利称之为异胞质;1889年荷兰学者德弗里斯称之为泛生子;1883年德国动物学家魏斯曼称之为种质。实际上魏斯曼所说的种质已经不再是单纯的臆测了,他已经指明生殖细胞的染色体便是种质,并且明确地区分种质和体质,认为种质可以影响体质,而体质不能影响种质,在理论上为遗传学的发展开辟了道路
孟德尔三大定律
从1910年到现在遗传学的发展大致可以分为三个时期:细胞遗传学时期、微生物遗传学时期和分子遗传学时期。

微生物遗传

遗传学
在这一时期中,采用微生物作为材料研究基因的原初作用、精细结构、化学本质、突变机制以及细菌的基因重组、基因调控等,取得了已往在高等动植物研究中难以取得的成果,从而丰富了遗传学的基础理论。1900~1910年人们只认识到孟德尔定律广泛适用于高等动植物,微生物遗传学时期的工作成就则使人们认识到遗传学的基本规律适用于包括人和噬菌体在内的一切生物。
分子遗传

遗传学
分子遗传学是在微生物遗传学和生物化学的基础上发展起来的。分子遗传学的基础研究工作都以微生物、特别是以大肠杆菌和它的噬菌体作为研究材料完成的;它的一些重要概念如基因和蛋白质的线性对应关系、基因调控等也都来自微生物遗传学的研究。分子遗传学在原核生物领域取得上述许多成就后,才逐渐在真核生物方面开展起来。
正像细胞遗传学研究推动了群体遗传学和进化遗传学的发展一样,分子遗传学也推动了其他遗传学分支学科的发展。遗传工程是在细菌质粒和噬苗体以及限制性内切酶研究的基础上发展起来的,它不但可以应用于工、农、医各个方面,而且还进一步推进分子遗传学和其他遗传学分支学科的研究。
基本内容

遗传学
一个受精卵通过有丝分裂而产生无数具有相同遗传组成的子细胞,它们怎样分化成为不同的组织是一个遗传学课题,有关这方面的研究属于发生遗传学。由一个受精卵产生的免疫恬性细胞能够分别产生各种不同的抗体球蛋白,这也是遗传学的一个课题,它的研究属于免疫遗传学。
遗传过程
基因表达
血红蛋白能够在哺乳动物血液中运输氧气。图中显示了血红蛋白在携氧和脱氧状态之间的结构变化。
单个氨基酸突变导致血红蛋白形成纤维。基因通常是通过生成所编码的蛋白质(执行细胞中大多数功能的复杂的生物大分子)来表现它们的功能性影响。蛋白质是由氨基酸所组成的线性链,而基因的DNA序列(通过RNA作为信息的中间载体)被用于产生特定的蛋白质的氨基酸序列。这一过程的第一步是由基因的DNA序列来生成一个序列互补的RNA分子,即基因的转录。
通过转录产生的RNA分子(信使RNA)被用于生产相应的氨基酸序列,这一转换过程被称为翻译。核酸序列中的每一组三个核苷酸组成一个密码子,可以被翻译为20种出现于蛋白质中的氨基酸中的一个,这种对应性被称为遗传密码。这种信息的传递是单一方向性的,即信息只能从核苷酸序列传递到氨基酸序列,而不能从氨基酸序列传递回核苷酸序列,这一现象被弗朗西斯·克里克称为分子生物学中心法则。
特定的氨基酸序列决定了对应蛋白质的独特的三维结构,而蛋白质结构则与它们的功能紧密相连。一些蛋白质是简单的结构分子,如形成纤维的胶原蛋白。蛋白质可以与其他蛋白质或小分子结合;例如,作为酶的蛋白质通过与底物分子结合来执行催化其化学反应的功能。蛋白质结构是动态的;例如,血红蛋白在哺乳动物血液中捕捉、运输和释放氧气分子的过程中能够发生微小的结构变化。
也有一些基因被转录为RNA分子后却不被翻译成蛋白质,这些RNA分子就被称为非编码RNA。在一些例子中,这些非编码RNA分子(如核糖体RNA和转运RNA)折叠形成结构并参与部分关键性细胞功能。还有的RNA(如microRNA)还能够通过与其他RNA分子杂交结合而发挥调控作用。基因序列上的单个核苷酸变化(密码子改变)可能会导致所编码蛋白质的氨基酸序列相应改变。由于蛋白质结构是由其氨基酸序列所决定的,一个氨基酸的变化就有可能通过使结构失去稳定性或改变蛋白质表面而影响与该蛋白质其他蛋白质和分子的相互作用,而引起蛋白质性质发生剧烈的改变。例如,镰刀型细胞贫血症是一种人类遗传性疾病,是由编码血红蛋白中的β-球蛋白亚基的基因中的一个核苷酸突变所引起的,这一突变导致一个氨基酸发生改变从而改变了血红蛋白的物理性质;在这一疾病中,突变的血红蛋白互相结合在一起,堆积而形成纤维,从而扭曲了携带血红蛋白的红血球的形状。这些扭曲的镰刀状细胞无法在血管中通畅地流动,容易堆积而阻塞血管或者被降解,从而引起贫血疾病。
先天后天
在人类遗传疾病苯丙酮尿症中,环境因素也具有重大的影响。导致苯丙酮尿症的突变破坏了机体降解苯丙氨酸的能力,导致具有毒性的中间产物分子在体内堆积,从而引起严重的进行性智能发育不全和癫痫。带有苯丙酮尿症突变的病人需要遵守严格的饮食,以避免摄入含苯丙氨酸的食品,才能保持正常的和健康的生活。暹罗猫具有温度敏感型突变,从而引起毛色的不同。虽然基因含有生物体所需功能的所有信息,环境依然在确定生物体最终的表现型中发挥着重要作用;这种两面性被称为“先天与后天”,也可以说,一个生物体的表现型依赖于遗传与环境的相互作用。这种相互作用的一个例子就是温度敏感型突变:蛋白质序列中的单个氨基酸突变通常不会改变该蛋白质的行为和与其他分子的相互作用关系,但却能够使该蛋白质结构变得不稳定。在一个高温环境中,分子的运动加快,分子间的碰撞也加强,这就使得这一蛋白质的结构被破坏从而无法发挥它的功能;而在一个低温环境中,蛋白质结构却可以保持稳定并能够发挥正常的功能。这类突变所引起的改变在暹罗猫毛色的变化中可以被观察到。这种猫体内一种负责生产色素的酶含有一个突变,这个突变能够导致这种酶在高温时变得不稳定并失去其功能。因此,在猫皮肤温度较低处(如四肢、尾部、面部等)的毛色为深色,而较高处为浅色。
基因调控
多细胞生物中的基因表达的差异性非常明显:虽然各类细胞都含有相同的基因组,却由于不同的基因表达而具有不同的结构和行为。多细胞生物中的所有细胞都来源于一个单一细胞,通过响应外部或细胞之间的信号而不断分化并逐渐建立不同的基因表达规律来产生不同的行为。因为没有一个单一基因能够负责多细胞生物中的各个组织的发育,因此这些规律应来自于许多细胞之间的复杂的相互作用。这些过程都要通过基因调控来完成。转录因子与DNA结合,影响了所结合基因的转录。一个生物体的基因组含有数千个基因,但并不是所有的基因都需要保持激活状态。基因的表达表现为被转录为mRNA,然后再被翻译成蛋白质;而细胞中存在许多方式可以来控制基因的表达,以便使蛋白质的产生符合细胞的需求。而控制基因表达“开关”的主要调控因子之一就是转录因子;它们是一类结合在基因的起始位点上的调控蛋白,可以激活或抑制基因的转录。例如,在大肠杆菌细菌基因组内存在着一系列合成色氨酸所需的基因。然而,当细菌细胞可以从环境中获得色氨酸时,这些基因就不被细胞所需要。色氨酸的存在直接影响了这些基因的活性,这是因为色氨酸分子会与色氨酸操纵子(一种转录因子)结合,引起操纵子结构变化,使得操纵子能够结合到合成色氨酸所需基因上。色氨酸操纵子阻断了这些基因的转录和表达,因而对色氨酸的合成进程产生了负反馈调控作用。
关系及应用
生物化学关系
遗传学与生物化学的关系最为密切,和其他许多生物学分支学科之间也有密切关系。例如发生遗传学和发育生物学之间的关系;行为遗传学同行为生物学之间的关系;生态遗传学同生态学之间的关系等。此外,遗传学和分类学之间也有着密切的关系,这不仅因为在分类学中应用了DNA碱基成分和染色体等作为指标,而且还因为物种的实质也必须从遗传学的角度去认识。
遗传学
40年代
40年代初,抗菌素工业的兴起推动了微生物遗传学的发展,微生物遗传学的发展又推动了抗菌素工业以及其他新兴的发酵工业的进步。随着微生物遗传学研究的深入,基因调控作用的原理被成功地应用到氨基酸等发酵工业中。此外杂交转导、转化等技术的采用也增加了育种的手段。
70年代
遗传学研究同人类本身密切相关。由于人类遗传学研究的开展,特别是应用体细胞遗传学和生化遗传学方法所取得的进展,对于遗传性疾病的种类和原因已经有很多了解;产前诊断和婴儿的遗传性疾病诊断已经逐渐推广;对于某些遗传性疾病的药物治疗也在研究中。免疫遗传学是组织移植和输血等医学实践的理论基础;药物遗传学和药物学有密切的关系;毒理遗传学关系到药物的安全使用和环境保护。用遗传工程技术对遗传性疾病进行基因治疗也正在进行探索。人类遗传学研究也是优生学的基础。
遗传学研究为致癌物质的检测提供了一系列的方法。虽然目前治疗癌症还没有十分有效的方法,但在环境污染日益严重的今天能够有效地检测环境中的致癌物质,便是一个重大的进展。癌症患病的倾向性是遗传的,癌症的起因又同DNA损伤修复有关,近年来癌基因的发现进一步说明癌症和遗传的密切关系,所以从长远观点来看,遗传学研究必将为全面控制癌症作出贡献。
近几年来,人类基因组研究的进展日新月异,而分子生物学技术也不断完善,随着基因组研究向各学科的不断渗透,这些学科的进展达到了前所未有的高度。在法医学上,STR位点和单核苷酸(SNP)位点检测分别是第二代、第三代DNA分析技术的核心,是继RFLPs(限制性片段长度多态性)VNTRs(可变数量串联重复序列多态性)研究而发展起来的检测技术。作为最前沿的刑事生物技术,DNA分析为法医物证检验提供了科学、可靠和快捷的手段,使物证鉴定从个体排除过渡到了可以作同一认定的水平,DNA检验能直接认定犯罪、为凶杀案、强奸杀人案、碎尸案、强奸致孕案等重大疑难案件的侦破提供准确可靠的依据。随着DNA技术的发展和应用,DNA标志系统的检测将成为破案的重要手段和途径。此方法作为亲子鉴定已经是非常成熟的,也是国际上公认的最好的一种方法。
新趋势
1991-1997年,中国曾邦哲[杰](Zeng BJ.)发表《结构论-泛进化理论》系列论文,阐述系统医药学(systems medicine)、系统生物工程(system biological engineering)与系统遗传学(system genetics)的概念,提出经典、分子与系统遗传学发展观,以及于2003年、2008年于国际遗传学大会,采用结构(structure)、系统(system)、图式(pattern)遗传学的词汇来描述系统科学方法、计算机技术研究生物系统遗传结构、生物系统形态图式之间的“基因型-表达型”复杂系统研究领域,以细胞信号传导、基因调控网路为核心研究细胞进化、细胞发育、细胞病理、细胞药理的细胞非线性系统动力学。
2003年挪威科学家称之为整合遗传学(integrative genetics)并建立了研究中心,2005年,国际上Cambien F.和 Laurence T.发表动脉硬化研究的系统遗传学观,Morahan G., Williams RW.等2007年(Bock G., Goode J. Eds.)论述系统遗传学将成为下一代遗传学。2005-2008年,国际系统遗传学飞速发展,欧美建立了许多系统遗传学研究中心和实验室。2008年在美国召开了整合与系统遗传学国际会议,2009年荷兰举办了系统遗传学国际会议,2008年美国国立卫生研究院(NIH)设立了肿瘤的系统遗传学研究专项基金。系统遗传学,采用计算机建模、系统数学方程、纳米高通量生物技术、微流控芯片实验等方法,研究基因组的结构逻辑、基因组精细结构进化、基因组稳定性、生物形态图式发生的细胞发生非线性系统动力学。
研究技术
一开始遗传学家们的研究对象很广泛,但逐渐地集中到一些特定物种(模式生物)的遗传学上。这是由于新的研究者更趋向于选择一些已经获得广泛研究的生物体作为研究目标,使得模式生物成为多数遗传学研究的基础。模式生物的遗传学研究包括基因调控以及发育和癌症相关基因的研究。
模式生物具有传代时间短、易于基因操纵等优点,使得它们成为流行的遗传学研究工具。目前广泛使用的模式生物包括:大肠杆菌(Escherichia coli)、酿酒酵母(Saccharomyces cerevisiae)、拟南芥(Arabidopsis thaliana)、线虫(Caenorhabditis elegans)、果蝇(Drosophila melanogaster)以及小鼠(Mus musculus)。
遗传学研究
医学遗传学的目的是了解基因变异与人类健康和疾病的关系。当寻找一个可能与某种疾病相关的未知基因时,研究者通常会用遗传连锁和遗传系谱来定位基因组上与该疾病相关的区域。在群体水平上,研究者会采用孟德尔随机法来寻找基因组上与该疾病相关的区域,这一方法也特别适用于不能被单个基因所定义的多基因性状。一旦候选基因被发现,就需要对模式生物中的对应基因(直系同源基因)进行更多的研究。对于遗传疾病的研究,越来越多发展起来的研究基因型的技术也被引入到药物遗传学中,来研究基因型如何影响药物反应。
癌症虽然不是传统意义上的遗传病,但被认为是一种遗传性疾病。癌症在机体内的产生过程是一个综合性事件。机体内的细胞在分裂过程中有一定几率会发生突变。这些突变虽然不会遗传给下一代,但会影响细胞的行为,在一些情况下会导致细胞更频繁地分裂。有许多生物学机制能够阻止这种情况的发生:信号被传递给这些不正常分裂的细胞并引发其死亡;但有时更多的突变使得细胞忽略这些信号。这时机体内的自然选择和逐渐积累起来的突变使得这些细胞开始无限制生长,从而成为癌症性肿瘤(恶性肿瘤),并侵染机体的各个器官。
研究技术
可以在实验室中对DNA进行操纵。限制性内切酶是一种常用的剪切特异性序列的酶,用于制造预定的DNA片断。然后利用DNA连接酶将这些片断重新连接,通过将不同来源地DNA片断连接到一起,就可以获得重组DNA。重组DNA技术通常被用于在质粒(一种短的环形DNA片断,含有少量基因)中,这常常与转基因生物的制造有关。将质粒转入细菌中,再在琼脂平板培养基上生长这些细菌(来分离菌落克隆),然后研究者们就可以用克隆菌落来扩增插入的质粒DNA片断(这一过程被称为分子克隆)。
DNA还能够通过一个被称为聚合酶链锁反应(又被称为PCR)的技术来进行扩增。利用特定的短的DNA序列,PCR技术可以分离和扩增DNA上的靶区域。因为只需要极少量的DNA就可以进行扩增,该技术也常常被用于DNA检测(检测特定DNA序列的存在与否)。
测序与基因组
DNA测序技术是遗传学研究中发展起来的一个最基本的技术,它使得研究者可以确定DNA片段的核苷酸序列。由弗雷德里克·桑格和他的同事于1977年发展出来的链终止测序法现在已经是DNA测序的常规手段。在这一技术的帮助下,研究者们能够对与人类疾病相关的DNA序列进行研究。
由于测序已经变得相对廉价,而且在计算机技术的辅助下,可以将大量不同片断的序列信息连接起来(这一过程被称为“基因组组装”),因此许多生物(包括人类)的基因组测序已经完成。这些技术也被用在测定人类基因组序列,使得人类基因组计划得以在2003年完成。随着新的高通量测序技术的发展,DNA测序的费用被大大降低,许多研究者希望能够将测定一个人的基因组信息的价格降到一千美元以内,从而使大众测序成为可能。
大量测定的基因组序列信息催生了一个新的研究领域——基因组学,研究者利用计算机软件查找和研究生物的全基因组中存在的规律。基因组学也能够被归类为生物信息学(利用计算的方法来分析生物学数据)下的一个领域。
基因异常
一个或多个基因异常,特别是隐性基因,是相当普遍的。每个人都携带有6~8个异常隐性基因。然而,这些基因并不引起细胞功能异常,除非有两个相似的隐性基因存在。一般人群中,具有两个相似隐性基因的个体机率非常小,但是在近亲婚配的孩子中,这种机率较高。在近亲婚配的群体中,比如基督教的阿曼门诺派,机率也较高。
一个人的基因组成称之为基因型。这些基因在人体存在的反映,即基因型的表达称之为表现型。
所有遗传特征(性状)为基因所编码。有一些特征,比如头发颜色、人与人之间的差异等,不能算是异常,然而,由异常基因表达产生的异常特性可能导致遗传疾病。
单基因异常
单个基因异常的影响取决于这个基因是显性还是隐性基因,和这个基因是否位于X染色体上(X连锁)。因为一个基因指导一种特定蛋白质的合成,因此异常基因就可能产生异常蛋白质或导致某种蛋白质含量异常,进而引起细胞功能的异常,最终导致机体形态或功能异常。
染色体遗传
位于常染色体上的异常显性基因产生的性状,可以导致畸形、疾病或有发展成某一疾病的倾向。
有这种性状的人,至少父母有一方具有同样性状,除非这种性状是由新的基因突变引起。
异常遗传性状常常是由新的遗传突变所致,而不是由父母遗传。
当父母一方具有异常性状,而另一方没有时,他们的每一个孩子有50%的机率遗传这种异常,有50%的机率没有遗传这种性状。如果父母之一具有两个这种异常基因(虽然很罕见),他们所有子女都将有异常性状。
不具异常性状的个体,即使他的兄弟姊妹有异常性状,因为他不携带这种基因,他的子女也不会遗传异常性状。
男性和女性受影响的可能性相同。
常常在每一代中都出现异常。
下列一些原则通常适用于由隐性基因决定的性状:
实际上,具有这种性状的人,其父母双方都可能携带有这种隐性基因,但父母双方均不表现出这种性状。
突变很少引起该性状的表达。
父母之一表现出性状,另一方携带隐性基因,没有表现性状,他们的子女中约有一半有可能表现出异常性状;而另一半将是隐性基因的携带者。假如另一方没有携带异常基因,他们的子女将无一人表现出异常性状,但他们所有的子女都将是携带者,并且可将这个异常基因遗传给他们子女的后代。
一个没有异常性状的人,如果兄弟姊妹有异常性状,他很可能携带有一个这种异常基因。
男性和女性受影响的可能性相同。
通常不是在每一代都有异常表现,除非父母双方均表现为异常。
引起严重疾病的显性基因是罕见的。因为这类基因趋于消失,带有引起严重疾病显性基因的人常常因为疾病太严重而没有生育能力。然而,有少数例外,如亨廷顿舞蹈病(见第67节),可引起严重的脑功能衰退,症状常开始于35岁之后,在症状表现之前,患者可能已经有了孩子。
隐性基因仅仅是在该个体具有两个同样的这种隐性基因时才有可能表达。仅有一个隐性基因的个体不表现出性状,但他是这种隐性基因的携带者,能够将此基因遗传给他的孩子。
X连锁基因
因为男性的Y染色体上的基因数目很少,在单个X染色体上的基因(X连锁基因)都是不成对的。因此,无论是显性基因或是隐性基因都能够表达。但是,女性有两条X染色体,适用于常染色体上基因的那些原则,同样适用于女性X连锁基因。除非成对的两个基因都是隐性的,通常只有显性基因才能表达。
假如一个异常X连锁基因是显性,受影响的男性会将异常基因遗传给他所有的女儿,但不会遗传给儿子。受影响的男性的儿子接受他的Y染色体,而Y染色体不携带异常基因。只带有一个异常基因的女性,可以把异常基因遗传给她的一半子女,儿子或女儿。
假如异常X连锁基因是隐性基因,几乎出现异常性状的均为男性。异常的男性只把异常基因传给女儿,她们都是携带者。携带者母亲并不表现异常,但可将异常基因遗传给一半儿子,这些儿子常常表现出异常性状。她们的女儿没有异常性状,但女儿中有一半是携带者。
红绿色盲,由X连锁隐性基因引起,男性发生率大约为10%,而女性则不常见。在男性,引起色盲的基因来自母亲,母亲是色盲患者或者是具有正常视力的色盲基因携带者。色盲基因不来自父亲,因为父亲提供Y染色体。色盲父亲的女儿极少是色盲,但常常是色盲基因的携带者。
等显性遗传,两个基因均表达。例如镰状细胞贫血;假如一个人有一个正常基因,同时又有一个异常基因,就可能产生正常和异常两种血红蛋白。
线粒体基因
基因遗传
癌细胞可能含有致癌基因,致癌基因是引起癌症的基因(也称肿瘤基因)(见第162节)。有时癌基因是出生前负责生长发育的基因的异常翻译,正常情况下,出生后它们就永远失活。如果这些致癌基因以后被再激活,可能导致癌症发生。它们是如何被再激活的尚不知道。
参考资料
[1]
经典遗传学与分子遗传学 · 大河网[引用日期2017-01-18]


