转录(遗传信息从DNA流向RNA的过程)

VLoG
次浏览
更新时间:2023-05-22
转录
遗传信息从DNA流向RNA的过程
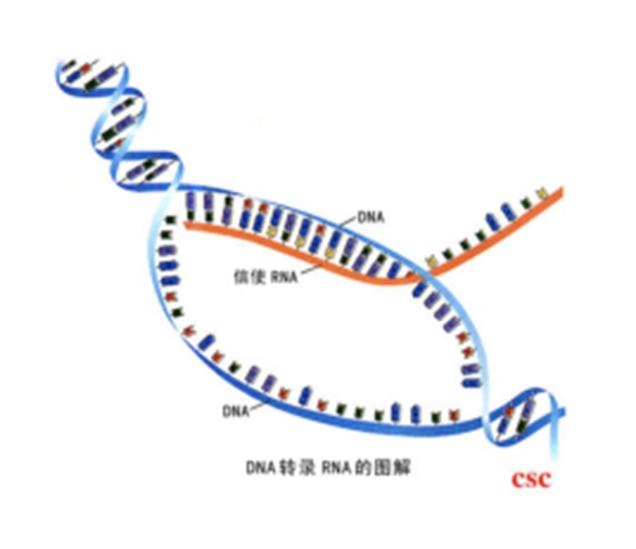
转录(Transcription)是遗传信息从DNA流向RNA的过程。即以双链DNA中的确定的一条链(模板链用于转录,编码链不用于转录)为模板,以A,U,C,G四种核糖核苷酸为原料,在RNA聚合酶催化下合成RNA的过程。作为蛋白质生物合成的第一步,进行转录时,一个基因会被读取并被复制为mRNA,即特定的DNA片断作为遗传信息模板,以依赖DNA的RNA聚合酶作为催化剂,通过碱基互补的原则合成前体mRNA。RNA聚合酶通过与一系列组分构成动态复合体,完成转录起始、延伸、终止等过程。生成的mRNA携有的密码子,进入核糖体后可以实现蛋白质的合成。转录仅以DNA的一条链作为模板,被选为模板的单链称为模板链,亦称无义链;另一条单链称为非模板链,即编码链,因编码链与转录生成的RNA序列T变为U外其他序列一致,所以又称有义链。DNA上的转录区域称为转录单位。
基本信息
| 中文名 | 转录 |
| 外文名 | Transcription |
| 含义 | 遗传信息由DNA转换到RNA的过程 |
| 意义 | 基因调节的主要阶段 |
定义
转录( transcription)是指以DNA为模板,以ATP、UTP、GTP和CTP为原料,按照碱基互补原则,在RNA聚合酶的作用下合成RNA的过程,是基因c表达的第一步。原核细胞只有一种RNA聚合酶;而真核细胞则有3种:RNA聚合酶I、RNA聚合酶Ⅱ及RNA聚合酶Ⅲ。DNA双链中作为转录模板的单链称为模板链( templatestrand)或反义链( antisense strand),另一条链则称为编码链( coding strand)或有义链( sense strand)(图2-6)。一个DNA分子上有许多基因,并非所有基因的编码区都在同一条单链上,因此模板链或编码链是相对某个基因的转录而言的。原核细胞的RNA转录:原核细胞的RNA聚合酶全酶(a2Bβ'σ)是由4条多肽链组成的核心酶加σ因子构成转录过程可划分为开始、延伸和终止三个阶段。①开始:σ因子识别DNA分子上的启动子并与之结合,将DNA双链局部解开,RNA合成开始,σ因子与核心酶分离。②延伸:RNA聚合酶沿模板链向前移动,使RNA链不断合成延长。③终止:原核细胞转录终止分依赖ρ因子和不依赖p因子两类。由于原核细胞没有核膜,且合成的RNA几乎不需进行复杂的加工修饰,故原核细胞的转录和翻译两个过程几乎可以同时进行。
转录过程
启动
RNA聚合酶正确识别DNA模板上的启动子并形成由酶、DNA和核苷三磷酸(NTP)构成的三元起始复合物(图1),转录即自此开始。DNA模板上的启动区域常含有TATAATG顺序,称普里布诺(Pribnow)盒或P盒。复合物中的核苷三磷酸一般为GTP,少数为ATP(见核苷酸),因而原始转录产物的5′端通常为三磷酸鸟苷(pppG)或腺苷三磷酸(pppA)。真核 DNA上的转录启动区域也有类似原核DNA的启动区结构,和在-30bp(即在酶和 DNA结合点的上游30核苷酸处,常以-30表示,bp为碱基对的简写)附近也含有TATA结构,称霍格内斯(Hogness)盒或 TATA盒。第一个核苷三磷酸与第二个核苷三磷酸缩合生成3′-5′磷酸二酯键后,则启动阶段结束,进入延伸阶段。
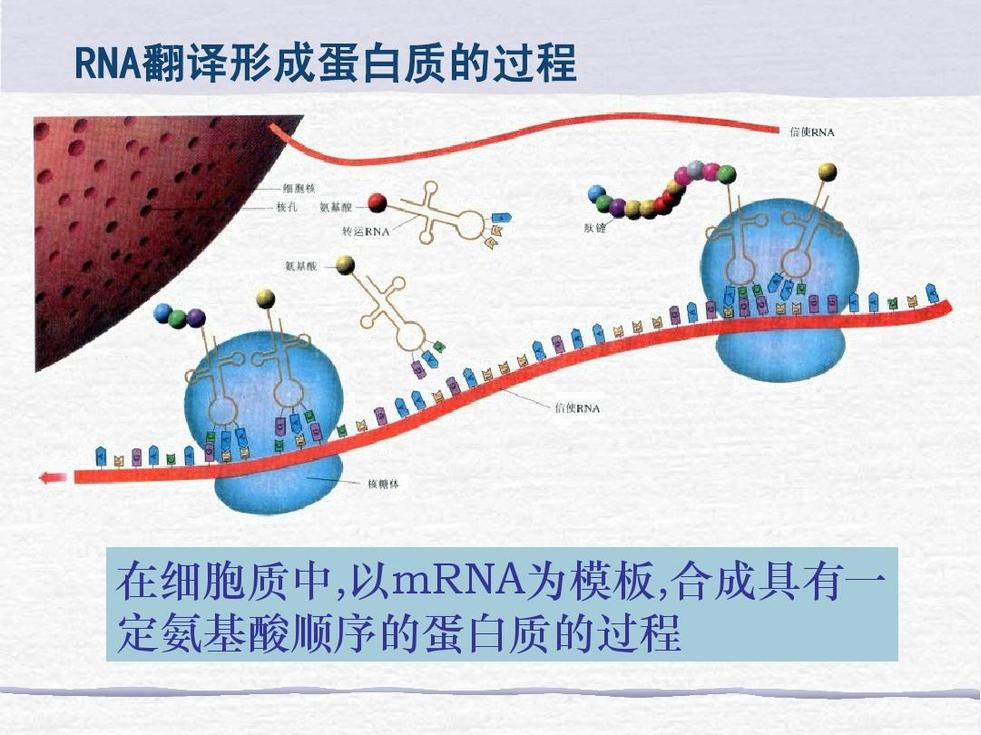
转录产物的后加工
mRNA前体的后加工
原核mRNA的原始转录产物(除个别噬菌体外)都可直接用于翻译,而真核mRNA一般都有相应的前体,前体必须经过后加工才能用于转译蛋白质。一般认为,真核mRNA的原始转录产物(也称原始转录前体), hn RNA(hetero-geneous nuclear RNA,核不均一RNA),最终被加工成成熟的mRNA(见信使核糖核酸)。
mRNA前体的后加工包括以下四方面(图2):①装上 5′端帽子:转录产物的 5′端通常要装上甲基化的帽子;有的转录产物 5′端有多余的顺序,则需切除后再装上帽子。②装上3′端多聚A尾巴:转录产物的3′端通常由多聚A聚合酶催化加上一段多聚A,多聚A尾巴的平均长度在20~200个核苷酸;有的转录产物的3′端有多余顺序,则需切除后再加上尾巴。装5′端帽子和3′端尾巴均可能在剪接之前就已完成。③剪接:将mRNA前体上的居间顺序切除,再将被隔开的蛋白质编码区连接起来。剪接过程是由细胞核小分子RNA(如U1RNA)参与完成的,被切除的居间顺序形成套索形(即lariat RNA中间体)。④修饰:mRNA分子内的某些部位常存在 N6 -甲基腺苷(见核苷酸),它是由甲基化酶催化产生的,也是在转录后加工时修饰的。

tRNA前体的后加工
目前分离得到的tRNA前体有两类:①含单个tRNA的tRNA前体,在5′端和3′端各有一段多余顺序;②含二个tRNA的tRNA前体,除5′端和3′端有长短不一的多余顺序外,在两个tRNA之间还有数目不等的核苷酸隔开。有的真核tRNA前体的反密码子环区含有一个居间顺序。
原核和真核生物tRNA前体的后加工有相似的步骤:①修饰:对tRNA分子上的部分核苷酸进行修饰(包括甲基化、酰化、硫代和重排等);②切除5′端和3′端多余核苷酸;③3′端不含CCA顺序的tRNA前体需装上CCA顺序(见转移核糖核酸)。原核与真核tRNA前体的加工过程还有不同的情况:①原核多顺反子tRNA前体,需加工时切开;②含有居间顺序的真核tRNA前体,加工时需除去居间顺序(图3)。
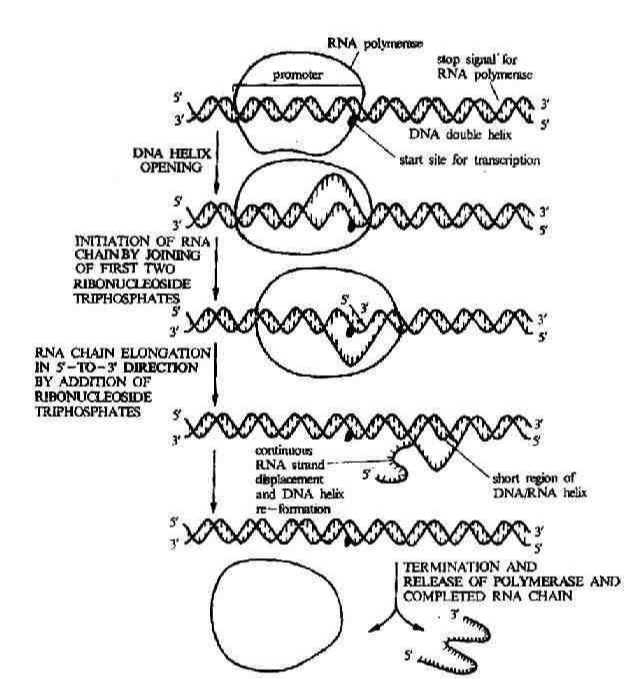
转录
rRNA前体的后加工 通常有如下步骤:①修饰:除5SrRNA外,rRNA分子上通常有修饰核苷酸(主要是甲基化核苷酸),它们都是在后加工时修饰的。一般认为核糖2′羟基的甲基化在碱基甲基化之前;②剪切:在rRNA前体分子的多余顺序处切开,产生许多中间前体,然后再切除中间前体末端的多余顺序(图4);③剪接:有的真核生物rRNA前体中存在有居间顺序的,须加工时除去。1982年T.R.切赫发现,在四膜虫(Tetrahymena)rRNA前体中,去除含有413个核苷酸的居间顺序是由rRNA前体自身催化完成的(图5)。在 5′-鸟苷酸的促进下经过自身催化作用将居间顺序切除,居间顺序前后的两个部分再连接起来,产生成熟的rRNA(5′-UpU-3′)和一个环状 RNA分子及一个15个核苷酸残基的小片段。rRNA前体的自身催化作用表明 RNA具有类似于酶的活性。这一发现突破了生物高分子中只有蛋白质才有催化作用的观念。同时对生物进化与生命起源等研究都将有重要的意义。
调节控制
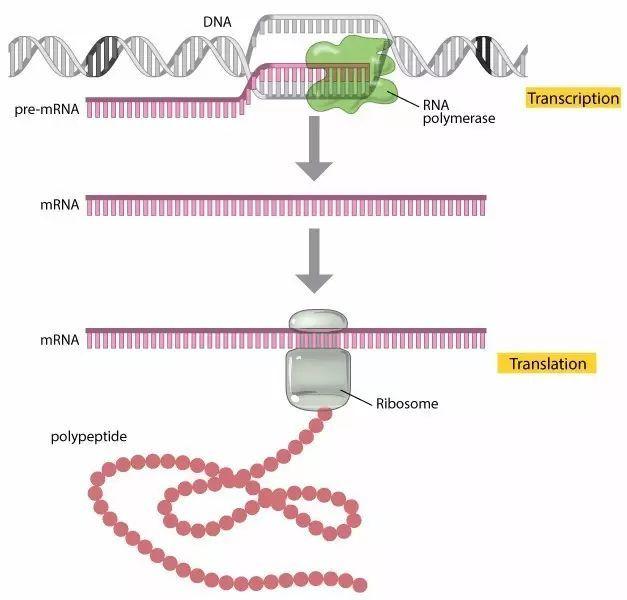
转录
对真核细胞基因转录的调节控制目前知道得很少。同种高等生物每个个体的各个体细胞都有全套相同的基因,只是由于在发育过程中基因表达的调节控制(包括转录的调节控制)不同,因而发育成各种不同的组织和器官。目前认为,动物(包括人)都含有癌基因,但有的致癌,有的则不致癌,这也可能是由于转录与翻译的调控不同。另外,真核DNA中的结构基因只占总量的10%左右,大部分 DNA顺序都可能起调节控制作用。真核生物也有诱导酶和诱导蛋白质,如干扰素就是由病毒或双链RNA等诱导产生的一种蛋白质。转录抑制剂 转录能被一些特异性的抑制剂抑制,有些抑制剂是治疗某些疾病的药物,有的则是研究转录机理的重要试剂。按照作用机理的不同,转录抑制剂分为两大类。第一类抑制剂特异性地与 DNA链结合,抑制模板的活性,使转录不能进行。这类抑制剂同时抑制DNA复制,例如:放线菌素D、纺锤菌素、远霉素、溴乙锭和黄曲霉素等。第二类抑制剂作用于RNA聚合酶,使RNA聚合酶的活性改变或丧失,从而抑制转录的进行。这类抑制剂只抑制转录,不影响复制,是研究转录机制和RNA聚合酶性质的重要工具,例如:利福平。曲张链霉素、利链霉素和α-鹅膏蕈碱等。
特点
转录中,一个基因会被读取并复制为mRNA。就是说,以特定的DNA片断作为模板,以DNA依赖的RNA合成酶作为催化剂,合成前体mRNA。
RNA聚合酶合成RNA时不需引物,但无校正功能。
举例
逆转录
转录过程
在转录过程中,DNA模板被转录方向是从3′端向5′端;RNA链的合成方向是从5′端向3′端。RNA的
合成一般分两步,第一步合成原始转录产物(过程包括转录的启动、延伸和终止);第二步转录产物的后加工,使无生物活性的原始转录产物转变成有生物功能的成熟RNA。但原核生物mRNA的原始转录产物一般不需后加工就能直接作为翻译蛋白质的模板。
区别
真核生物RNA的转录与原核生物RNA的转录过程在总体上基本相同,但是,其过程要复杂得多,主要有以下几点不同:
⒈真核生物RNA的转录是在细胞核内进行的,而蛋白质的合成则是在细胞质内进行的。所以,RNA转录后首先必须从核内运输到细胞质内,才能指导蛋白质的合成。
⒉真核生物一个mRNA分子一般只含有一个基因,原核生物的一个mRNA分子通常含有多个基因,而除少数较低等真核生物外,一个mRNA分子一般只含有一个基因,编码一条多肽链。
⒊真核生物RNA聚合酶较多,在原核生物中只有一种RNA聚合酶,催化所有RNA的合成,而在真核生物中则有RNA聚合酶Ⅰ、RNA聚合酶Ⅱ和RNA聚合酶Ⅲ三种不同酶,分别催化不同种类型RNA的合成。三种RNA聚合酶都是由10个以上亚基组成的复合酶。RNA聚合酶Ⅰ存在于细胞核内,催化合成除5SrRNA以外的所有rRNA的合成;RNA聚合酶Ⅱ催化合成mRNA前体,即不均一核RNA(hnRNA)的合成;RNA聚合酶Ⅲ催化tRNA和小核RNA的合成。
⒋真核生物RNA聚合酶不能独立转录RNA。原核生物中RNA聚合酶可以直接起始转录合成RNA,真核生物则不能。在真核生物中,三种RNA聚合酶都必须在蛋白质转录因子的协助下才能进行RNA的转录。另外,RNA聚合酶对转录启动子的识别,也比原核生物更加复杂,如对RNA聚合酶Ⅱ来说,至少有三个DNA的保守序列与其转录的起始有关,第一个称为TATA框(TATAbox),具有共有序列TATAAAA,其位置在转录起始点的上游约为25个核苷酸处,它的作用可能与原核生物中的-10共有序列相似,与转录起始位置的确定有关。第二个共有序列称为CCAAT框(CCAATbox),具有共有序列GGAACCTCT,位于转录起始位置上游约为50-500个核苷酸处。如果该序列缺失会极大地降低生物的活体转录水平。第三个区域一般称为增强子(enhancer),其位置可以在转录起始位置的上游,也可以在下游或者在基因之内。它虽不直接与转录复合体结合,但可以显著提高转录效率。
抑制剂
转录能被一些特异性的抑制剂抑制,有些抑制剂是治疗某些疾病的药物,有的则是研究转录机理的重要试剂。按照作用机理的不同,转录抑制剂分为两大类。第一类抑制剂特异性地与DNA链结合,抑制模板的活性,使转录不能进行。这类抑制剂同时抑制DNA复制,例如:放线菌素D、纺锤菌素、远霉素、溴乙锭和黄曲霉素等。第二类抑制剂作用于RNA聚合酶,使RNA聚合酶的活性改变或丧失,从而抑制转录的进行。这类抑制剂只抑制转录,不影响复制,是研究转录机制和RNA聚合酶性质的重要工具,例如:利福平等。
解旋酶
在真核细胞中,RNA聚合酶通常不能单独发挥转录作用,而需要与其他转录因子共同协作,有些辅助功能的转录因子就是解旋酶。
以反式作用影响转录的因子可统称为转录因子(transcriptionfactors,TF)。RNA聚合酶是一种反式作用于转录的蛋白因子。与RNA聚合酶Ⅰ、Ⅱ、Ⅲ相应的转录因子分别称为TFⅠ、TFⅡ、TFⅢ,对TFⅡ研究最多。表19-2列出真核基因转录需要基本的TFⅡ。?
以前认为与TATA盒结合的蛋白因子是TFⅡ-D,后来发现TFⅡ-D实际包括两类成分:与TATA盒结合的蛋白是TBP(TATAboxbindingprotein),是唯一能识别TATA盒并与其结合的转录因子,是三种RNA聚合酶转录时都需要的;其他称为TBP相关因子(TBP?associatedfactorsTAF),至少包括8种能与TBP紧密结合的因子。转录前先是TFⅡ-D与TATA盒结合;继而TFⅡ-B以其C端与TBP-DNA复合体结合,其N端则能与RNA聚合酶Ⅱ亲和结合,接着由两个亚基组成的TFⅡ-F加入装配,TFⅡ-F能与RNA聚合酶形成复合体,还具有依赖于ATP供给能量的DNA解旋酶活性,能解开前方的DNA双螺旋,在转录链延伸中起作用。这样,启动子序列就与TFⅡ-D、B、F及RNA聚合酶Ⅱ结合形成一个“最低限度”能有转录功能基础的转录前起始复合物(pre?intitiationcomplex,PIC),能转录mRNA。TFⅡ-H是多亚基蛋白复合体,具有依赖于ATP供给能量的DNA解旋酶活性,在转录链延伸中发挥作用;TFⅡ-E是两个亚基组成的四聚体,不直接与DNA结合而可能是与TFⅡ-B联系,能提高ATP酶的活性;TFⅡ-E和TFⅡ-H的加入就形成完整的转录复合体,能转录延伸生成长链RNA,TFⅡ-A能稳定TFⅡ-D与TATA盒的结合,提高转录效率,但不是转录复合体一定需要的。

解旋酶


