防御素(富含二硫键的阳离子型多肽)

VLoG
次浏览
更新时间:2023-05-19
基本信息
| 中文名 | 防御素 |
| 外文名 | Defensin |
| 活性 | 免疫调控,抗菌 |
| 分布 | 真菌 、植物与动物 |
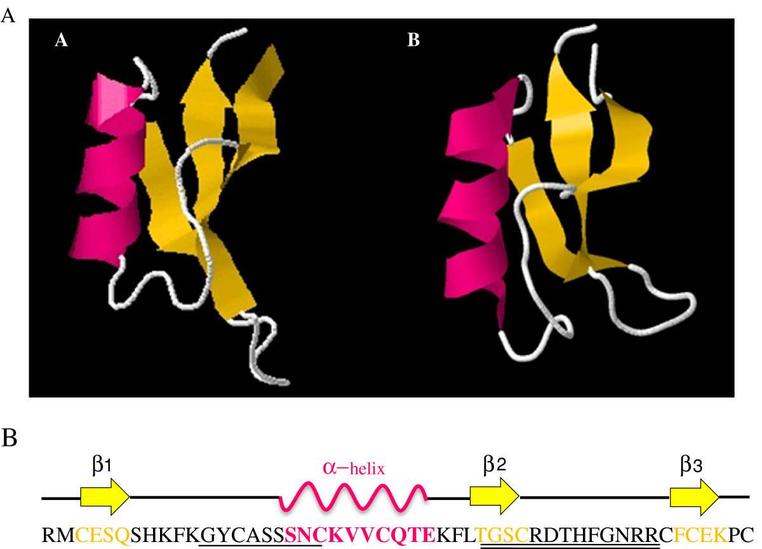
| 二级结构 | αβ/全β |
防御素介绍
天然的防御素分布在从植物、低等动物到哺乳动物等几乎所有的生物类群中,是生物长期与疾病斗争中进化而来的,也是其自身防御体系的重要组成部分。大部分防御素和其他蛋白质类物质一样,由基因直接编码,可通过基因工程技术开发生产。
研究背景
1966年,美国科学家Zeya和Spitznagel首次在哺乳动物老鼠和豚鼠的多形核嗜中性白细胞中发现一类具有抗菌活性的碱性多肽,将其称之为“溶酶体阳离子蛋白”。这就是后来人们称之为防御素的物质。到目前,人们已经鉴定了四百余种防御素。
1985年,美国加州大学Robert Lehrer博士首次命名防御素(defensins),并报道了兔防御素MCP-1 及MCP-2对流感病毒和单纯疱疹病毒具有直接中和灭活作用。次年,该研究组还报道了人体内三种alpha防御素 (Humanα-defensin或human neutrophil peptide, HNP)对流感病毒、巨细胞病毒、水泡性口炎病毒等均有中和作用。
2002年,美国洛克菲勒大学艾伦·戴蒙德艾滋病研究中心的华人科学家张林琦在《SCIENCE》杂志上发表文章,报道人体内抑制艾滋病病毒复制的物质是T淋巴细胞分泌的α-防御素,这个发现为艾滋病的控制与治疗提供了新的思路与方向,引起生物医学界的广泛关注。
2003年开始,和仕康公司的核心研发人员在构建海洋生物海马等物种的cDNA文库时发现了大量防御素基因,并分离到五种天然防御素。2007年开始,他们依托南海海洋生物技术国家工程中心等科研转化平台将防御素发酵制剂作为饲料添加剂进行大量动物实验,结果表明该制剂可显著提升动物免疫力和防病抗病水平。
防御素的分类
(一)哺乳动物类防御素
α-防御素是于1980年美国Lehrer实验室从兔肺巨噬细胞中首先分离得到的、阳离子性极强的小分子抗菌肽,称为defensin,后被归为α-防御素;主要分布于人、兔、猪、鼠类的嗜中性粒细胞、兔子的齿槽巨嗜细胞及人类和啮齿动物的小肠潘氏细胞。α-防御素分子链内二硫键的连接位置分别为Cys1-Cys6、Cys2-Cys4、Cys3-Cys5。其中Cys1-Cys6连接N端和C端,形成分子大环。
β-防御素是Diamond等(1991)首先在牛的气管黏膜上皮细胞中发现的,后又在牛粒性白细胞中发现了13 种与其序列高度相似, 但其共有序与α-防御素不同,故被命名为β-防御素;主要分布于牛的骨髓和人及多种动物(牛、羊、猪、骆驼、驯鹿、小鼠、大鼠)的胃肠道、呼吸道、舌、牙龈、肾、皮肤的上皮中,近来在梅花鹿舌粘膜的上皮细胞内也发现了该短肽。单核细胞和巨噬细胞通常缺乏防御素,但是他们可以释放诱导上皮细胞合成β-防御素的信使。β-防御素分子链内二硫键的连接位置分别为Cys1-Cys5、Cys2- Cys4、Cys3-Cys6。
θ-防御素是于2002年Trabi等用反向高效液相层析色谱法从猕猴的白细胞中分离出来的1种成环状结构分子,又被称为猕猴θ型防御素-1(RTD-1),主要分布于巨嗜细胞内。θ-防御素的结构不同于α和β-防御素,其前体(已经发现3种)是α-防御素类似物,由1个终止密码子从α-防御素3个半胱氨酸碳骨架的第4个残基处截断,从截短的α-防御素前体切掉1个 9个氨基酸 的片段,然后从头到尾进行剪切,直到其他的相同或相似的九肽出现。成熟的θ-防御素是2个半防御素的修饰与结合的产物,它的前体(称之为半防御素)是变异的α-防御素基因和1个未成熟的终止密码子的编码产物,导致每1个前体仅仅包含3个半胱氨酸残基。θ-防御素分子链内二硫键的连接位置分别为Cys1-Cys4、Cys2-Cys5、Cys3-Cys6,连接形成环状结构。
(二)昆虫类防御素
在1988年由Masturyama在1种半翅目昆虫肉蝇中发现了第1 种昆虫防御素。后来Dimarcq等先从果蝇中分离到1种病原菌诱导的抗菌肽,发现果蝇的防御素基因是单拷贝且无内含子,据此推测昆虫的防御素是独立进化而来的。后来经研究发现,昆虫防御素与哺乳动物防御素在二硫键的连接方式以及三维空间构型上是截然不同的,从而证明了昆虫防御素和哺乳动物防御素不具同源性。绝大多数昆虫的防御素带1个净正电荷,分子量为 4ku 左右,由34~51个氨基酸组成,氨基酸序列中都含有6个保守半胱氨酸, 3个分子内二硫键形成β片状结构和1个α螺旋结构,二硫键以Cys1-Cys4、Cys2-Cys5、Cys3-Cys6连接。
(三)植物类防御素
在1990年由Mendez等从小麦、大麦种子首先分离得到植物防御素r1-P和r1-H,用NMR光谱法测定其三维结构,发现与哺乳动物和昆虫防御素相似。最近Thevissen也通过比较植物和昆虫防御素与真菌的葡糖苷鞘氨醇的相互作用,得知它们是由同一个前体细胞祖先进化而来的。植物防御素的分子质量小于5ku,由45~54个氨基酸组成, 3个分子内二硫键形成1个反向的β片状结构和1个α螺旋结构,含有4 对二硫键且连接方式为Cys1-Cys8、Cys2-Cys5、Cys3-Cys6、Cys4-Cys7。
生物活性
(一)抗菌作用
防御素能有效杀灭革兰阴性细菌和革兰阳性细菌。在体外浓度为 10~100 mg/L 的防御素即对多种细菌具有杀伤作用,而防御素在中性粒细胞中的浓度为 g/L 级,远远超过上述数值,这表明在体内防御素可能具有更强的杀菌活性,目前研究发现防御素对革兰阳性细菌的杀伤能力明显要强于革兰阴性细菌。在体外,HBD-2 对大肠杆菌的半数致死量(LD50)为 0.46nmol/ml,最小抑菌浓度(MIC)为 15 μg/ml,而对绿脓杆菌、金黄色葡萄球菌的 MIC 为62 μg/ml。体外实验证明大多数防御素的 MIC 范围为0.5-10 μmol/L。
对于防御素抗菌活性机制,大部分研究者认为主要和微生物的细胞膜结构有关。防御素发挥抗菌作用可分为三个阶段:
(3)内容物外泄。通道形成后,防御素进入细胞内的同时,其他胞外分子也伴随进入(如肽、蛋白质或无机离子),而靶细胞的重要物质(如盐离子和大分子)渗出,致使靶细胞发生不可逆损伤而死亡。
(二)抗病毒作用
防御素能杀灭一些被膜病毒,如 HIV、疱疹病毒、水泡型口炎病毒,但对无衣壳病毒却无效。θ-防御素还具有抗滤过性病原体和抗毒素作用。体内实验显示,防御素可延缓或根除兔梅毒病,并使兔牙周炎龈下菌群恢复正常。防御素主要通过与病毒外壳蛋白结合从而导致病毒丧失生物活性,这一特殊作用机理也使得微生物不易对其产生抵抗性。防御素可直接抑制病毒,对病毒的抑制程度依赖于防御素浓度以及分子内二硫键的紧密程度,其抗病毒功效同样受时间、pH 值、离子强度和温度等因素的影响,在中性及低离子强度条件下,防御素具有较强的抗病毒活性,而在实验体系中加入血清或血清蛋白则可大大削弱防御素的抗病毒功效。有关防御素的抗病毒作用机理,大体上可以总结为如下三点:
1、闭门羹---- 阻止病毒入侵宿主细胞
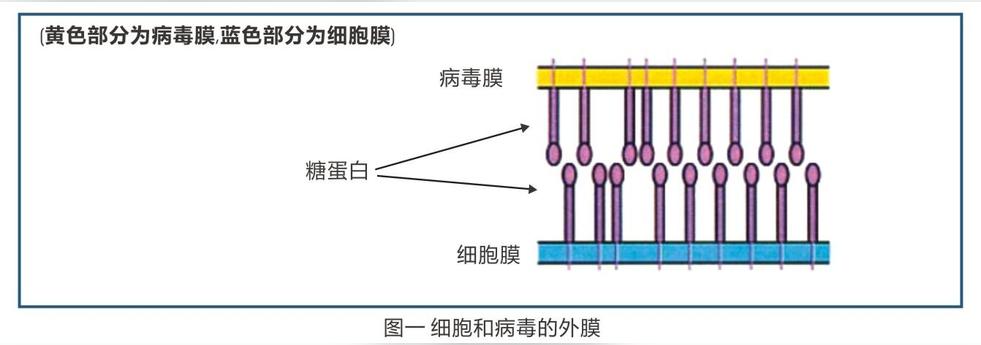
防御素则斜插在糖蛋白上以防病毒向细胞糖蛋白的扩散(见图二),让病毒吃个“闭门羹”无法进入细胞。没能进入细胞的病毒接着会被免疫系统的细胞摧毁。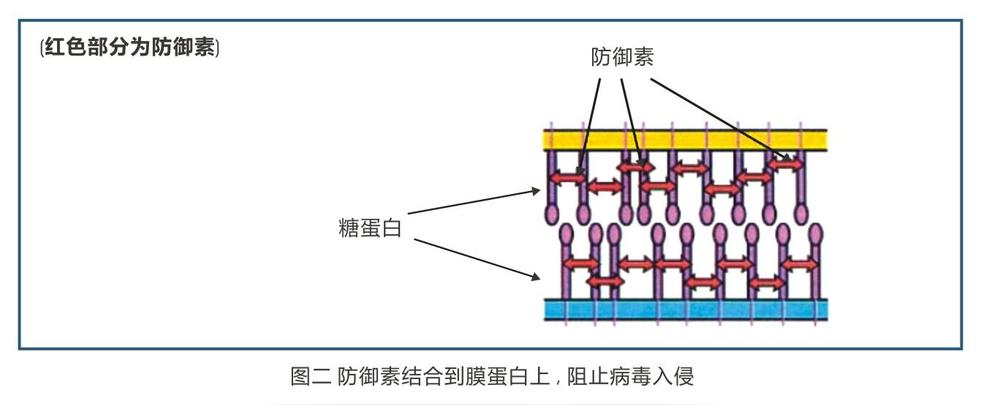

防御素
2、突破口----杀灭病毒
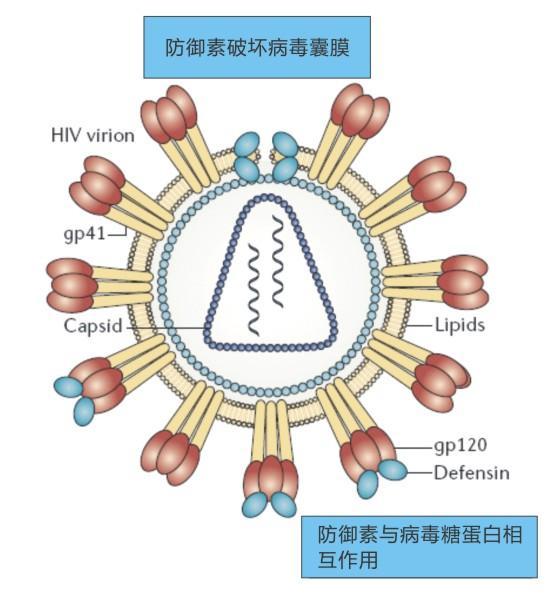
万一病毒进入细胞内,防御素可以与细胞膜表面促肾上腺皮质激素 (ACTH),人硫酸肝素糖蛋白(HSPG),低密度脂蛋白受体(LDLR)等相结合,从而启动G蛋白偶联型受体的级联放大反应,进一步激活磷酸激酶C。这些细胞信使作用就像埋伏在胞内的地雷,可以阻止整合到宿主基因组之前的病毒复合体进入细胞核或者阻止病毒的基因转录。整合不到宿主基因组的病毒随后被摧毁。
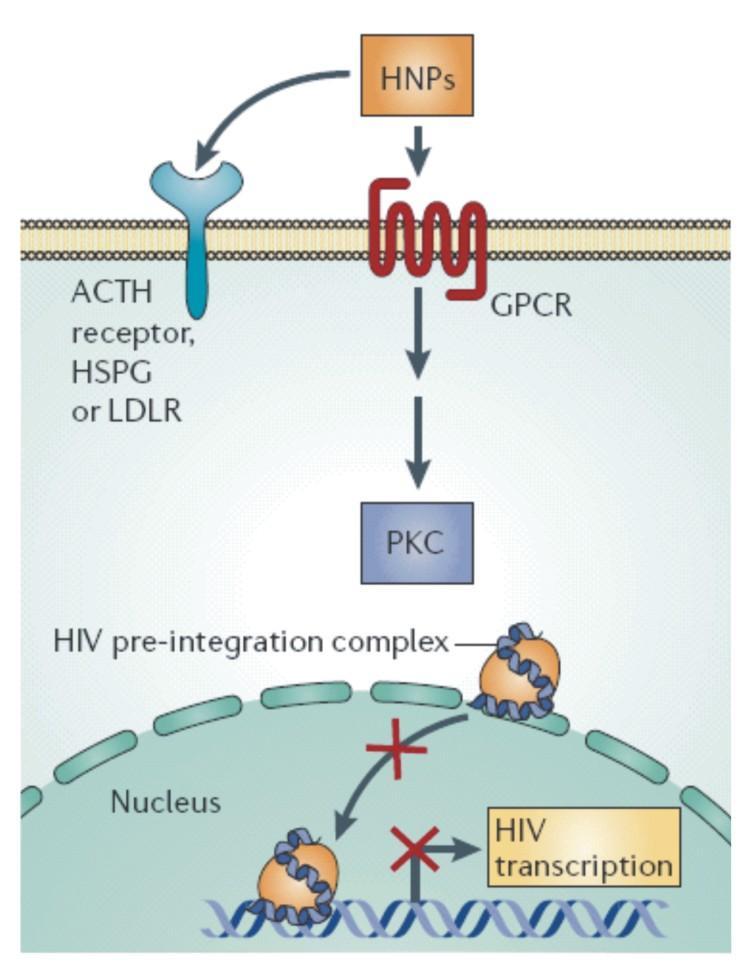
防御素
防御素对正常和恶性哺乳动物细胞都表现出非特异性的细胞毒作用,并且对人淋巴细胞及实体瘤细胞的细胞毒作用更为显著,尤其对抗 TNF 的 U9TR 细胞、抗NK细胞细胞毒因子的小鼠淋巴瘤 YAC-1 细胞和人组织细胞淋巴瘤 U937 细胞具有杀伤性。在体外,HNP可抑制烟酰胺腺嘌呤二核苷酸(NADPH)氧化酶的活性,HNP的细胞毒活性部分依赖于靶细胞膜脂质成分的代谢活性,它还可以与过氧化氢酶协同诱导细胞毒作用。体外观察纯化防御素对人及小鼠数种肿瘤细胞的杀伤作用,发现防御素对白血病细胞系、淋巴瘤细胞系及实体瘤细胞均具有细胞毒作用,并且这种抗肿瘤功效与其作用时间和剂量呈正相关。
(四)免疫调节作用
防御素不仅可以直接抵抗病原微生物,而且还具有免疫调节作用。防御素通过细胞信号传递的作用,增强非特异性免疫细胞,尤其是巨噬细胞的活性和趋化性。防御素还可以促进机体T细胞的趋化和增殖,增强机体免疫应答能力,调节特异性免疫,增强生物机体主动防御功能。
防御素能够作为一种效应分子激活巨噬细胞、DC、气管上皮细胞等细胞表面受体从而启动获得性免疫系统,并将先天性免疫和获得性免疫有机连接。现已证明一些 α-防御素、β-防御素对 T 细胞、单核细胞以及未成熟的 DC 具有趋化活性,可诱导单核细胞和上皮细胞产生细胞因子。
人、鼠、猪、兔的中性粒细胞防御素可以诱导肥大细胞脱颗粒并释放组胺。β-防御素还能通过与人趋化因子受体6(CCR6)结合,从而吸引不成熟的树突状细胞(DC)和记忆T细胞(Tm)至炎症部位,激活细胞免疫和体液免疫。此外,防御素还可以直接促进感染部位中性粒细胞的补充和积聚。
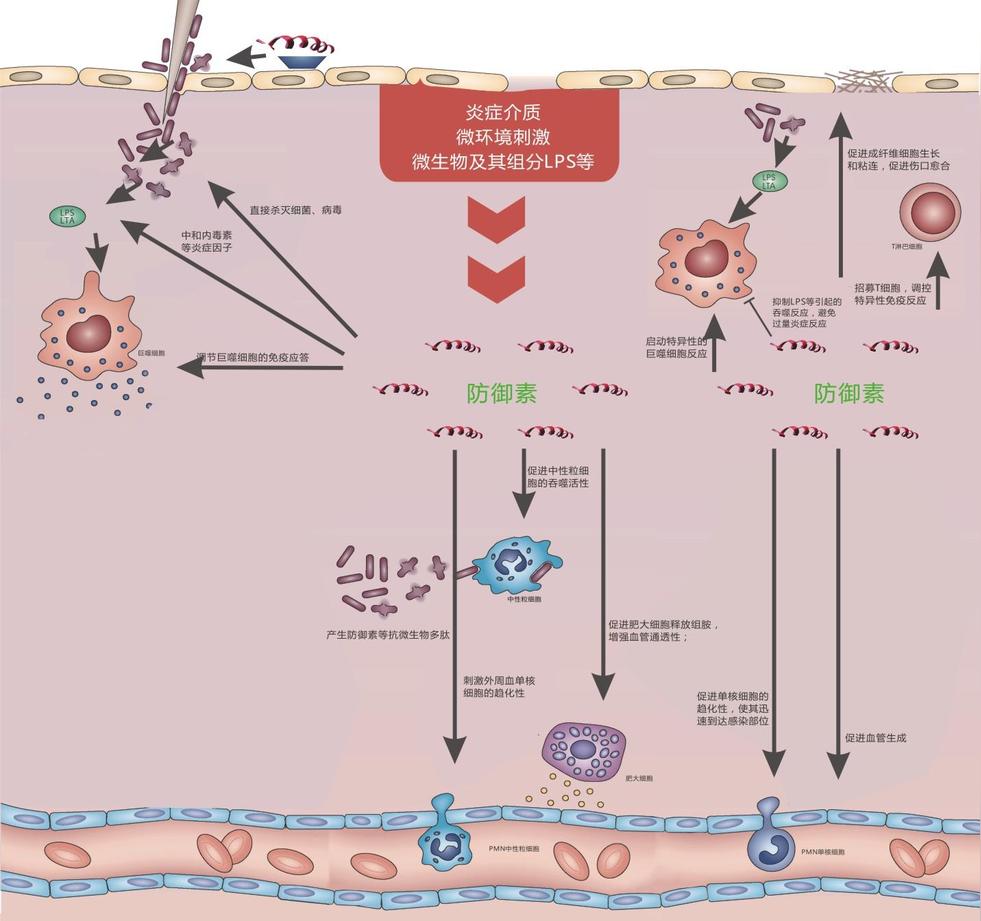
防御素
防御素的应用
(一)在医学中的应用
1、防御素与口腔黏膜疾病的关系
作为机体天然免疫的重要组成部分,防御素家族在口腔黏膜宿主防御及免疫应答过程中发挥着重要作用,与多种口腔黏膜疾病(口腔溃疡、扁平苔藓、白斑及口腔念珠菌病等)关系密切。现今关于这方面的研究尚处于起步阶段,主要以体外实验为主,其作用机制也多为推测,而有关防御素在体内的具体作用、有无毒性或其他不良反应等方面的研究甚少,还需进一步全面多层次的研究探讨,以阐明防御素在口腔黏膜疾病发生发展过程中的具体作用及其机制,为口腔黏膜疾病防治提供新的策略。
2、防御素与炎症性肠病的关系
克罗恩病(CD)是一种严重的肠道慢性炎症性疾病,病变部位可发生于消化道的任何部位,病变形式多样,其发病是遗传因素与环境因素共同作用的结果。肠道防御素是机体抵御外源性致病因子的重要屏障,而克罗恩病患者肠道防御素表达水平却存在着较大差异。回肠段病变的克罗恩病患者,回肠潘氏细胞防御素HD-5、HD-6 的表达较正常人和非回肠段病变克罗恩病患者明显降低,这种变化与 NOD2(nucleotide oligomerization domain)基因突变密切相关;结肠组织活检发现无 NOD2 基因突变的克罗恩病患者结肠组织HD-5、HD-6 的表达水平反而增高,而 HD-5、HD-6 水平降低将影响肠道黏膜的天然屏障作用,导致黏膜细菌附着引发进一步损害。
以往认为溃疡性结肠炎(UC)在欧美国家比较多见,但近年来中国文献报道的病例数也显著增多。UC 病变主要累及结肠,正常人结肠黏膜很少表达 HD-5 和 HD-6,而UC 患者结肠上皮细胞中 HD-5 和 HD-6 mRNA 的表达水平显著高于正常人,且 HD-6 的表达还具有特异性,即在非感染性的结肠炎组织中表达并不明显。HBD-1 在肠道上皮细胞呈持续性表达,且不受外界因子影响,具有基础防御功能。但应用实时 PCR 技术检测发现 UC 和克罗恩病结肠黏膜组织 HBD-1 基因表达水平均有所降低,这将可能促使肠道黏膜抗菌活性下降,从而导致细菌入侵机体引发炎症反应。在 UC 患者的病变结肠组织中,HBD-2 可被诱导性表达,而 HBD-3 的表达水平和 HBD-2 存在一定的相关性,其在 UC 时表达亦有明显增加。此外,HBD-4 mRNA的表达水平在 UC 肠上皮细胞中也明显增高。
慢性阻塞性肺部疾病(COPD)是一种以气道受限为特征的慢性气道炎症性疾病,患病率及病死率均较高,其中呼吸道感染是COPD 发病及加剧的一个重要因素。HBD-2 与呼吸系统疾病关系紧密,在肺部黏膜防御中具有重要作用。研究证实,HBD-2 的产生或作用被抑制后,机体将发生肺部感染,据此推测人为补充外源性防御素可能会阻止细菌的定植和感染的发生。已有研究证实,重组 HBD-2 对肺损伤具有保护作用,可减少绿脓杆菌感染大鼠肺组织的菌落数。COPD 患者存在气道、肺实质和肺血管炎症,而肺上皮细胞中高表达的 HBD-2 能通过对肺部炎症因子的调节,阻止全身性炎症反应(SIRS)甚至多器官功能衰竭(MODS)的发生发展。此外,重组 HBD-2 还可调控脓毒症诱发的肺组织细胞的过度凋亡,从而减轻肺组织损伤,保护肺功能。
4、防御素的临床药用研究
防御素抗菌谱广泛,作用机理特殊,微生物不宜产生抗药性,因此具有很大的药用潜力。2008年,全球首个防御素药物(PMX-30063)在加拿大完成一期临床试验,其对十余种耐药菌效果显著。针对金黄色葡萄球菌引起的急性细菌性皮肤感染这一适应症目前已经完成Ⅱ期临床研究。它通过直接靶向细菌细胞膜而达到杀菌的目的,不产生耐药性,从而有望解决日渐增多的细菌耐药性问题。
(二)在食品工业中的应用
(三)在畜牧业中的应用
抗生素添加剂的使用严重破坏了动物肠道的微生态平衡,药物残留也影响了畜产品的品质和人类健康,而来源于哺乳动物的防御素相对分子质量较小,热稳定性和水溶性均较好,可以在肠道内吸收。由于防御素属于多肽成分,在体内容易被蛋白酶降解为氨基酸,动物采食后在体内一般无残留。利用基因工程方法生产环保型防御素饲料添加剂或通过日粮添加,都可调控防御素基因的表达并实现其功效,如减少死胎、木乃伊及残疾;提高初生重及仔猪出生均匀度;提高仔猪断奶重、断奶成活率,降低哺乳期仔猪发病率;降低哺乳母猪发病率,减少母、仔猪各种应激;改善母猪亚健康和繁殖障碍,延长母猪使用年限;提高保育猪成活率,降低发病率;提高保育猪日增重、降低料肉比;改善免疫抑制,提高免疫应答能力,抗体水平更整齐。
(四)在农业中的应用
基因工程制备


